Сравнительная физиология динозавров и птиц. Популярно о малоизвестном. Часть 1 «Кости титанов»

В одном из моих предыдущих обзоров, вы ознакомились с хронологией крупнейших геологических и космических катастроф в истории нашей планеты. Последний из 5 крупнейших «апокалипсисов», предположительно поставил точку на более чем 150 миллионах лет существования «царей» фауны мезозойской эры — динозаврах. Впрочем «ужасные ящеры» все же оставили после себя палеонтологические «следы», позволяющие нам подробно ознакомиться с нелегкой жизнью этих рептилий.
В нескольких частях этого объемного обзора, я попытаюсь изложить вам малоизвестные подробности о физиологии и жизнедеятельности древних гигантских (и не очень) «ужасных» рептилий. Ниже в общих чертах будет описана сравнительная характеристика скелета, интеллекта и генетики динозавров с таковыми у их современных потомков — птиц, и родственных им крокодилов.
«Непохожие родственники»
В целом фиксируется огромное различие в строении скелета современных птиц и динозавров, объясняется это 160 миллионами лет адаптации определенной группы манирапторов к активному полету, вызвавшее кардинальную «перестройку» скелета и физиологии птиц. Нечто подобное наблюдается и у млекопитающих при сравнении копытных животных и их морских родственников (китообразных).
Подобное обретение существенных различий в морфологии родственных животных при их адаптации к разным экологическим нишам в науке именуется дивергенцией.
Кто кому родственник? Систематика хищных динозавров и птицМанирапторы являются большой группой динозавров, включающих в себя как современных птиц, так и их близких родственников — дейнонихозавров (небольшие хищники с «ужасным когтем»). Сами дейнонихозавры делились на троодонтид (по названию самого известного рода — троодона, жившего в конце эпохи динозавров), и дромеозавров (напр. бамбираптор и дейноних). В свою очередь манирапторы, вместе с тиранозавридами и орнитомимозаврами («имитаторами птиц»), входили в еще более многочисленную группу тираннорапторов. Последние вместе с комсогнатами (мелкие хищные тероподы) образовывали самую многочисленную группу теропод — целлурозавров.Наиболее изученные представители семейства дейнонихозавров, перьевой покров предполагается по следам «перьевых» ячеек сохранившихся на крылоподобных кистях велоцираптора и дейнониха
Однако целлурозавры не охватывают всех известных хищников теропод. В юрском (и раннем меловом) периоде доминировали в основном карнозавры, которые разошлись (генетически) с целлурозаврами ок. 170 млн лет назад, и с которыми входят в гигантский инфраотряд тетануров. Из наиболее значимых карнозавров следует отметить кархарадонтозавра, гиганотозаврва и аллозаврид. Спинозавр, крупнейший хищный теропод эры динозавров, тоже входил в группу титануров, но как представитель отдельного от целлурозавров отряда спинозаврид.
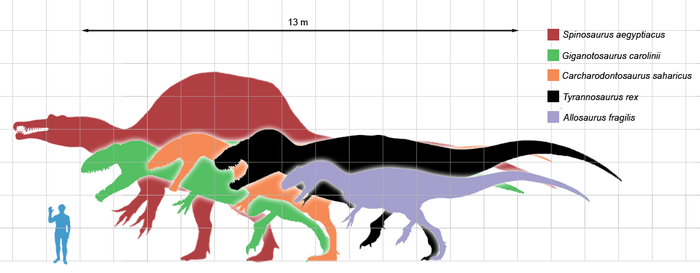
Относительные размеры крупнейших хищных теропод
Кроме тетануров известны и другие группы хищников. Один из главных «злодеев» видео игры «Турок» — карнотавр, относился к инфраотряду цератозаврид, относительно примитивных небольших хищных динозавров.

Антонимом дивергенции является конвергенция — процесс обретения схожих морфологических черт разными группами животных, при их адаптации к схожим условиям жизнедеятельности. Наглядный пример конвергенции на примере (слева на право) земноводной афанераммы, пресмыкающегося крокодила и млекопитающего амбулоцета (предка китообразных)
В общих чертах морфология динозавров представляла из себя нечто среднее между архаичными чертами пресмыкающихся (строение костей, генетика и метаболизм) и прогрессивными чертами птиц и млекопитающих (прямохождение, перьевой покров, строение челюстного аппарата травоядных динозавров). Доминирование тех или иных (прогрессивных или архаичных) черт скорее всего сильно варьировалось у разных отрядов и даже семейств динозавров.
Всех динозавров делят на две основные группы — ящеротазовых и птицетазовых:
• У двуногих ящеротазовых динозавров — теропод (схема ниже) центр тяжести располагался на линии таза, тяжелый хвост уравновешивал массивную голову животного. Лобковая (красный цвет) и седалищная (зеленый) кости таза располагались перпендикулярно к позвоночнику вдоль задних конечностей животного. Синим цветом выделена подвздошная кость, на которую крепятся мышцы бедра (в т.ч. и у человека).
Тираннозавр рекс, типичный ящеротазовый хищный теропод, родственный птицам по группе целлурозавров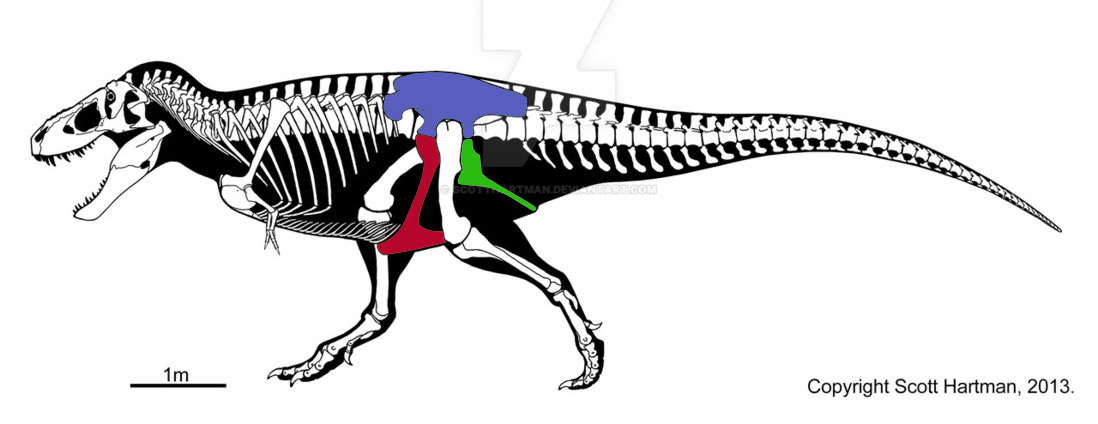
• Тазобедренный сустав основной части «четвероногих» растительноядных динозавров значительно отличался от такового у теропод. В частности лобковая и седалищная кость были расположены позади таза животных параллельно друг другу (аналогично птицам, так называемые «птицетазовые динозавры»). Впрочем, такое разделение на «двуногих» и «четвероногих» весьма условно, так четвероногие зауроподы имели схожий с тероподами тазобедренный сустав, а многие птицетазовые динозавры активно практиковали передвижение на задних конечностях.
Птицетазовый динозавр семейства гадрозавров («утконосые» динозавры)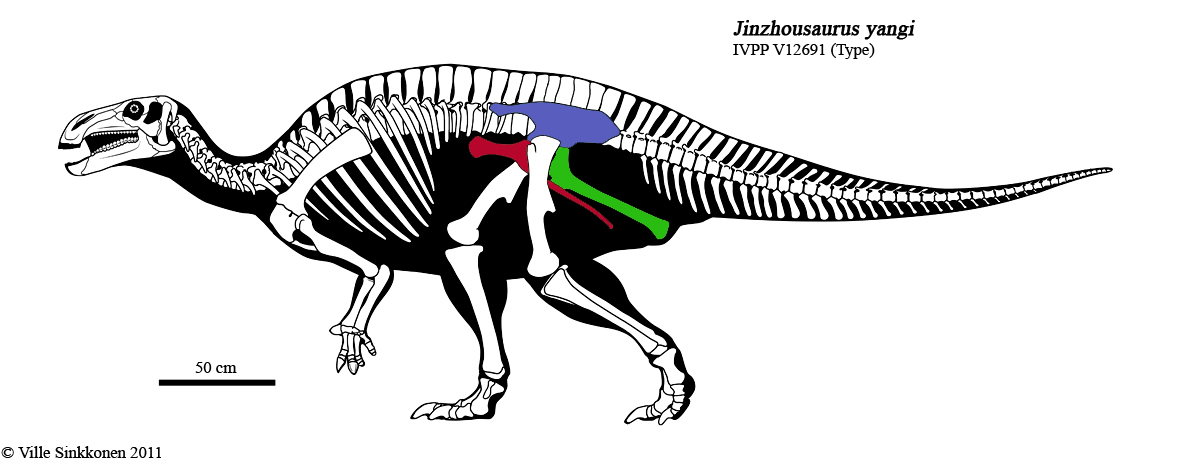
Зауропод из рода брахиозавров, можно отметить его принадлежность к ящеротазовым динозаврам, несмотря на передвижение на четырех конечностях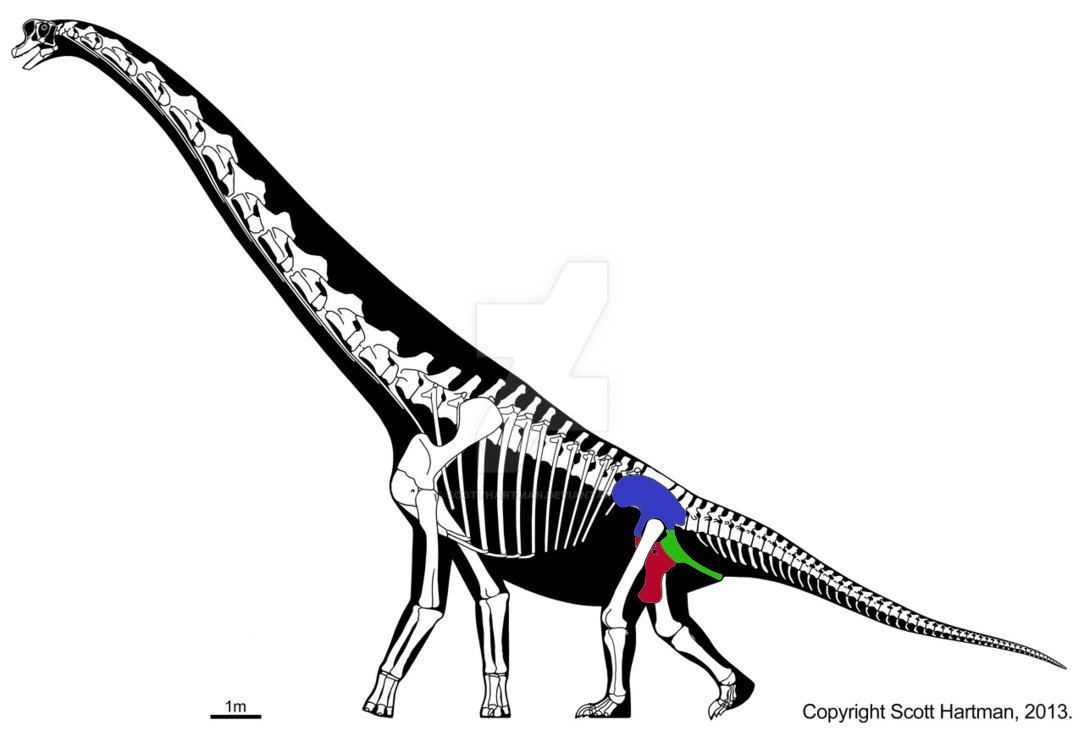
Тазобедренный сустав птиц значительно менее мобилен, лобковая и седалищная кости направлены назад и сращены друг с другом и нередко, с задней частью таза и хвостовых позвонков, образуя сложный крестец. Хвост птиц сильно редуцирован в пигостиль (который имелся и у некоторых овираптозавров), позвоночник малоподвижен. Ребра птиц имеют крючковатые отростки, позволяющие грудному отделу животного не двигаться при полете. На концах ребер большинства птиц крепится грудина и киль, на которые в свою очередь крепятся сильные маховые мышцы крыльев. Ребра и позвоночник динозавров были более мобильными чем у птиц, киля они так же не имели (впрочем, нет его и у современных не летающих птиц типа страусов).
Отдельно следует упомянуть разное строение шейных позвонков птиц и динозавров. У последних они амфицельные (двояковогнутые) или процельные/опистоцельные (вогнуты с одной стороны и выпуклы с другой, как у большинства пресмыкающихся). Шейные позвонки у птиц гетероцельные — седлообразные, выпукло-вогнутые с обеих сторон, что обеспечивает им большую подвижность шейного отдела, недоступную ни одному известному виду динозавров и млекопитающих.
Скелет современной птицы (голубя)
1 — череп, 2 — шейные позвонки, 3 — вилочка 4 — клювовидный отросток, 5 — крючковидные отростки ребер, 6 — киль, 7 — коленная чашечка, 8 — цевка, 9 — пальцы, 10 — большая берцовая кость (тибиотарсус), 11 — малая берцовая кость (тибиотарсус) 12 — бедро, 13 — седалищная кость, 14 — лобковая кость, 15 — подвздошная кость, 16 — хвостовые позвонки 17 — пигостиль, 18 — сложный крестец, 19 — лопатка, 20 — поясничные позвонки, 21 — плечевая кость, 22 — локтевая кость, 23 — лучевая кость, 24 — запястья, 25 — кисть, 26 — пальцы, 27 — крылышко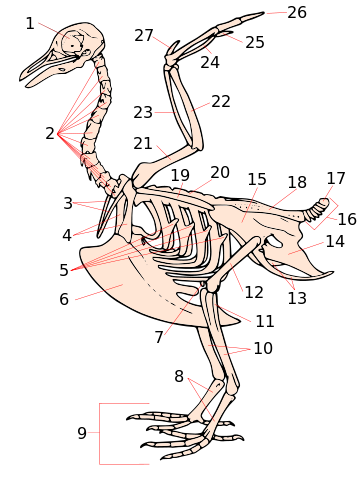
Бедренные кости птиц, даже не летающих, малоподвижны и направленны в сторону друг от друга. Перемещаются сухопутные птицы преимущественно с помощью движения тибиотарзуса (голень птицы с приросшейся малой берцовой костью) и цевки (сросшиеся кости плюсны). Т.е. тибиотарзус птиц, механически, выполняет роль бедренной кости у млекопитающих и динозавров. Конечности динозавров, напротив, располагались параллельно друг другу и под телом животных, в противоположность остальным пресмыкающимся и аналогично современным млекопитающим. Очевидно динозавры не были пресмыкающимися в буквальном смысле этого слова. Голень динозавров, как и голень млекопитающих, состояла из двух берцовых костей, многие (особенно маленькие динозавры) не имели цевки, и аналогично человеку, имели не сросшиеся кости плюсны.
Слева на право, сравнение задних конечностей теропода семейста тиранозаврид (вида рекс), волчьих млекопитающих (лисицы) и сухопутных птиц (страус). 1- кость бедра, 2 — кости голени (большая и малая берцовые/ тибиарзус у птиц), 3 — кости плюсны, сросшиеся у птиц в цевку (за исключением пингвинов) и некоторых динозавров — теропод. Зеленым цветом обозначены кости активно участвующие в передвижении животного. Грубо говоря, роль колена у современных птиц выполняет голеностопный сустав
Моделирование бега динозавров (галлимимуса и тиранозавра) из Парка Юрского периода, в сравнении с бегом современного страуса (внизу)
Малоподвижные бедра птиц позволяют им активно перемещаться по земле, не рискуя схлопыванием воздушных мешков, необходимых птицам для активного дыхания (в полете и беге легкие птицы не расширяются как у млекопитающих, воздух через них «прокачивают» те же воздушные мешки). Некоторые палеонтологи принимают этот факт как косвенное свидетельство отличной от птичьей дыхательной системы у теропод.
Обнаружение аэростеона, хищного теропода группы карнозавров, жившего 80 млн лет назад, отчасти опровергает выводы выше. «Воздушные кости» динозавра предположительно несут следы присутствия воздушных мешков. Изначально такие мешки могли служить для терморегуляции динозавров, и уже потом, с совершенствованием метаболизма начали использоваться для интенсивного газообмена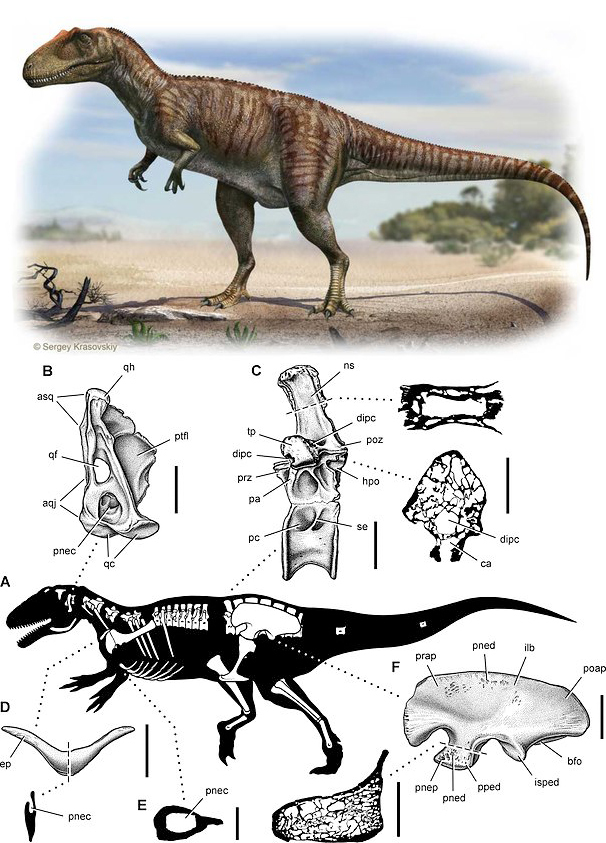
Еще сильнее различия между двумя группами животных проявляются при сравнительном анализе их черепов. Череп птицы крепится к шейным позвонкам снизу, шейные позвонки же динозавров крепились сзади черепа. Череп птиц имеет шарообразную форму и имеет две диапсидные дуги, верхняя из которых сильно редуцированна. Череп динозавров менее сферичен и обладает двумя развитыми диапсидными дугами (чаще всего с височными отверстиями). Челюсти динозавров устроены сложно, и подобно млекопитающим заполнены зубами. Современные птицы лишены зубов, и вместо сложного челюстного аппарата обладают относительно простым, легким, но прочным клювом.
Оговорка. Последнее сравнение не совсем корректно, так как ученым известны как «беззубые» динозавры (овирапторы), так и «зубастые» птицы (энансиорнисовые птицы, господствовавшие в меловой период, имели динозавроподобный челюстной аппарат.)
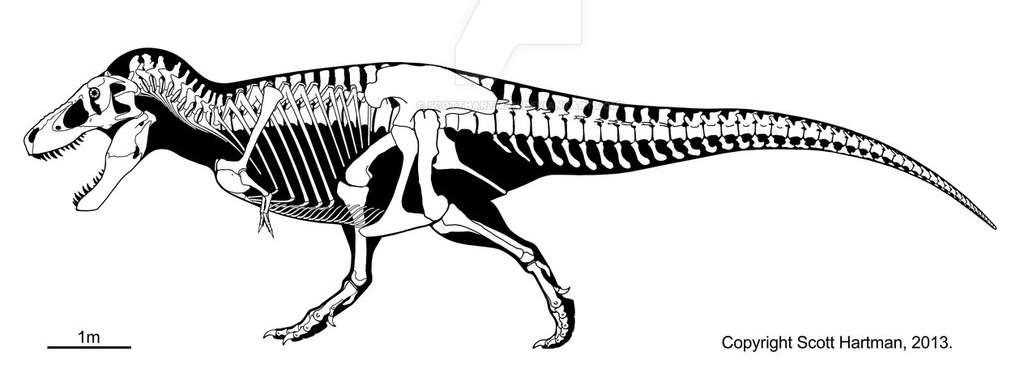
Графическое представление эволюции прогрессивных черт динозавров от «ужасных ящеров» и до «ужасных птиц»(Временные масштабы и порядок появления животных не соблюдены)Тираннозавр Рекс, живший 70–65 млн лет назад, представлял из себя весьма типичного (если не сказать эталонного) теропода. Из типичных черт динозавра в глаза бросаются две мощные задние конечности, ящеротазовый тазобедренных сустав, и массивный хвост, служащий балансиром для крупной и грузной головы животного. Череп прикреплен к шейным позвонкам сзади, имеются мощные челюсти покрытые зубами, пред глазничные отверстия (облегчающие череп) и за глазничные диапсидные отверстия куда крепились челюстные мышцы животного (аналог височной впадины человека). На фоне более близких родственников птиц — дромеозавров, выделяется отсутствием вилочки (срощенные кости ключицы) и сильно редуцированными относительными размерами передних конечностей
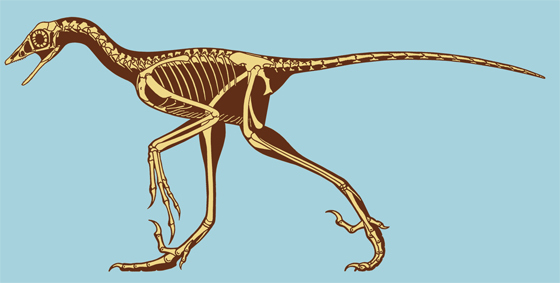
Дейноних, живший на 40 млн лет раньше тираннозавра рекс, обладал столь похожими на птиц чертами своего скелета (при жизни животное было размером с волка), что послужил для палеонтологов (особенности для знаменитого Джона Острома), одним из первых убедительных свидетельств происхождения птиц от динозавров. В частности кисти его передних конечностей очень сильно напоминают кисти крыльев первоптицы-археоптерикса, и отдаленно кисти современных птиц. Будучи «изломанными», кисти предположительно складывались на боках животного и скорее всего обладали перьями (как и у птицеподобных предков дейнониха). На ребрах имеются крючки как и у птиц. Легкие кости животного и жесткий, покрытый сухожилиями хвост (улучшающий маневренность при поворотах), указывал на его высокую скорость, и впервые навел палеонтологов на мысль о теплокровности динозавров. Как и все дромеозавры, дейноних имел «вилочку» в передней части груди, служащей для современных птиц «амортизатором» при сильных взмахах крыльев
Археоптерикс, живший 150 млн лет назад, впервые показавший прямую связь между птицами и пресмыкающимися, и сам считающийся одной из первых птиц, в целом сильно напоминает уменьшенного до размеров голубя дейнониха. Отличается от последнего большими относительными размерами мозга, уменьшением размеров челюстей и редукцией диапсидного отверстия. Археоптерикс не был предком птиц, но считается ближайшим его родственником, имел развитое оперение, но летать не умел, планируя с дерева на дерево
Микрораптор, динозавр из группы дромеозавров размером с археоптерикса, так же очень сильно напоминал своего более древнего родственника, что делает их скелеты практически неотличимыми для не специалистов. Животное жило 125 млн. лет назад и по образу жизни напоминало археоптерикса, обладая развитым перьевым покровом черного цвета. Охотился на насекомых и ранних птиц, которых глотал практически целиком. Как и выше упомянутый дромеозавр дейноних, лобковые и седалищные кости животного имеют тенденцию «загибания» назад
Современник микрораптора, и его «сосед» по региону (северо восточный Китай, южная Монголия), конфуциорнис уже находился на полпути между динозаврами и птицами. Его челюсти были лишены зубов, плечевой сустав был сильно развит, а вместо длинного хвоста птица уже имела пигостиль — место крепления хвостового оперения, улучшавшего маневренность при полете. Как и у микрораптора, на ребрах птицы имелись крючки, жестко фиксировавшие брюшную полость при полете. Место сочленения шейных позвонков и черепа находилось на полпути между его положением у динозавров и современных птиц. Однако строение плечевого сустава не позволяло птице полноценно освоить полет, ровно как и недоразвитый киль. Тазобедренный сустав все еще сильно напоминал таковой у дромеозавров
Синорнисы были одними из первых птиц, освоившими полноценный полет. Несмотря на это механика их полета сильно отличалась от таковой у современных птиц, как и у остальных энансиорнисовых птиц («противоптиц»). Имелся киль, пигостиль, а пальцы передних конечностей частично были сращены друг с другом. Челюстной аппарат и тазобедренный сустав больше был похож на таковой у динозавров, нежели современных (веерохвостных) птиц. Практический полный скелет синорниса был обнаружен в брюхе ископаемого микрораптора, который на них и охотился
Микрораптор, нападающий на группу синорнисов
Впрочем, древнейшими пернатыми, освоившими полноценный полет были предки современных нам веерохвостых птиц (Archaeornithura meemannae), «кузены» энантиорнитов по кладе Орнитоторасов — эуорниты. Последние отделились от энантиорнисов предположительно в самом конце юрского периода 140–145 млн. лет назад. Уже тогда начала проявляться тенденция специализации эуорнитов к охоте на мелководье и на береговых линиях. Позднемеловые ихтиорнисообразные вели схожий с чайками и крачками образ жизни и физиологически практически не отличались от современных птиц (смотрите изображение внизу), за исключением наличия зубов на обеих челюстях. Из колоссального разнообразия планирующих и летающих динозавров, мел-палеогенное вымирание пережила лишь небольшая часть эуорнитов, предков современных птиц (неорнитов).
«Ужасные птицы»
После мел-палеогенного вымирания птицы, одновременно с млекопитающими начали заполнять экологические ниши, оставленные их собратьями динозаврами. Так, уже через 15 млн лет появились первые гигантские сухопутные птицы — не летающие журавлеобразные. Размерами (до 2,5 м в высоту при весе до 300 кг) и образом жизни они мало отличались от своих родственников дромеозавров, так же будучи в основном хищными животными. Адаптация бесхвостой птицы к сухопутному образу жизни отразилась в смещении центра тяжести в центр тела животного. Упростил «работу» природе и облегченный череп птицы, вместо сложной челюсти вооруженный легким, но прочным клювом. В итоге современные сухопутные птицы представляют из себя более рационализированные версии динозавров-теропод, без затрат веса на мощные мышцы хвоста и шеи. Гигантские журавлеобразные господствовали в Америке последние 50 млн лет, проиграв в борьбе за выживание хищным млекопитающим (волчьи и кошачьи) около 400 000 лет назад. Ниже схема скелета крупнейшего журавлеобразного группы фороракосовых — Келенкена.Момент охоты другого представителя группы форорракосовых, по имени которого она и названна — фороракоса. Размеры хищника сильно преувеличены, на самом деле фороракосы были лишь немногим выше взрослого человека (до 2 м), но весили вдвое больше и действительно были грозными охотниками. Хотя фороракосы и жили одновременно с родом Homo, однако обитали исключительно в американской части света и с людьми не сталкивались
Подробнее о «ужасных птицах» в моем будущем обзоре «Затерянный мир Южной Америки».

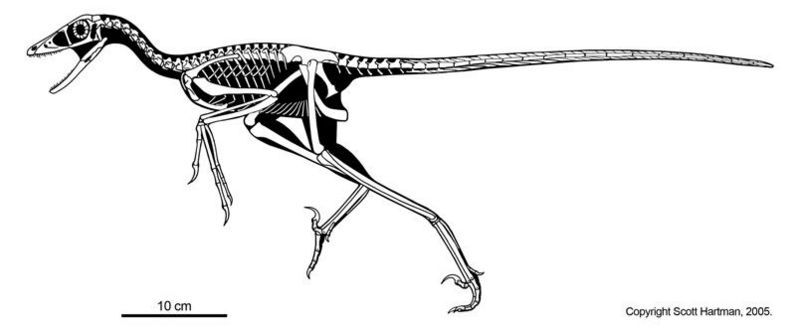
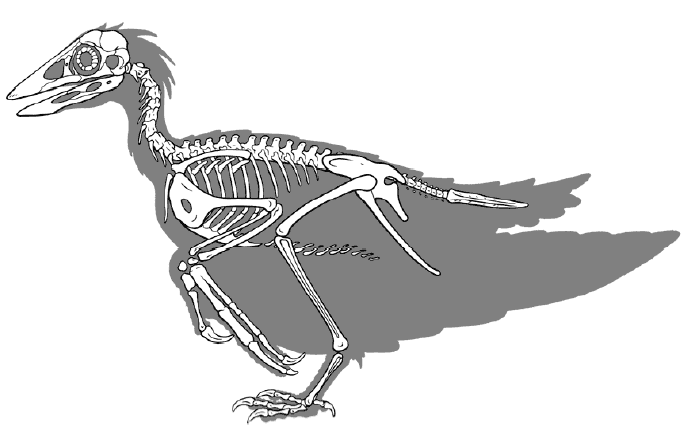
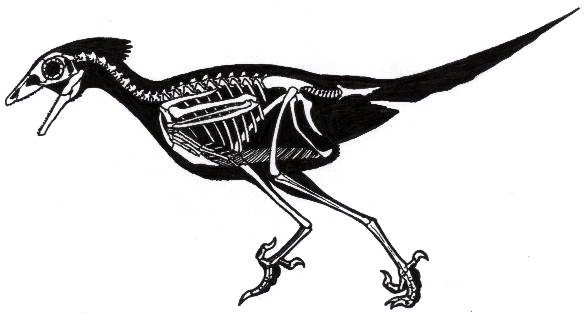

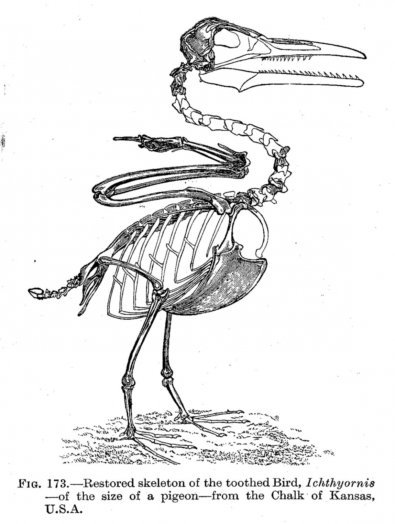
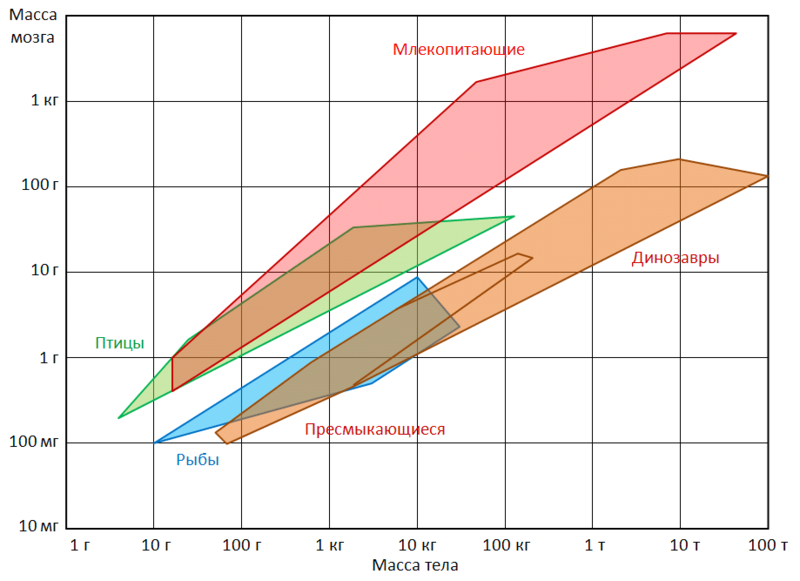
Относительные размеры мозга птиц (Коэффициент энцифализации) так же чаще всего, больше чем у динозавров аналогичного размера.
Коэффициент энцефализации упрощенно выражает отношение мозга животного к размерам его организма. Косвенно оно выражает уровень организации центральной нервной системы — интеллекта животного. КЭ не всегда объективно представляет истинный уровень интеллекта даже внутри одного вида, однако в целом, с эволюционной точки зрения, имеет линейную зависимость от уровня организации живых существ. Так самые высокие значения КЭ имеют высшие птицы и млекопитающие (примерно паритетные). Уже среди данной группы животных наиболее сложное поведение наблюдают у животных с наивысшими значениями КЭ (приматы и китообразные у млекопитающих и врановые с попугаеобразными у птиц).
Данный метод позволяет нам представить уровень интеллекта типичный для динозавров позднего мела.
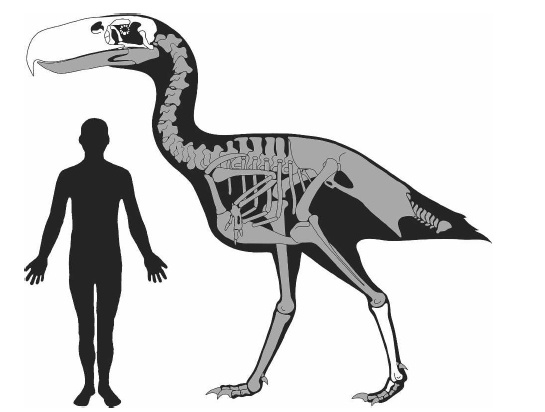
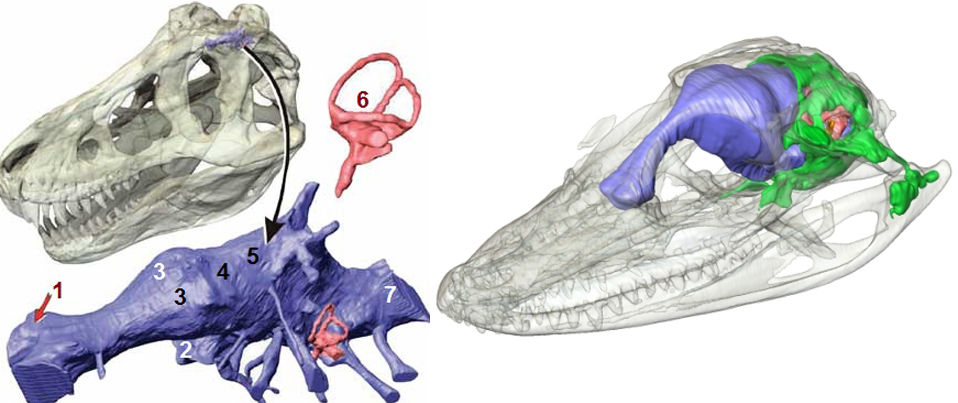
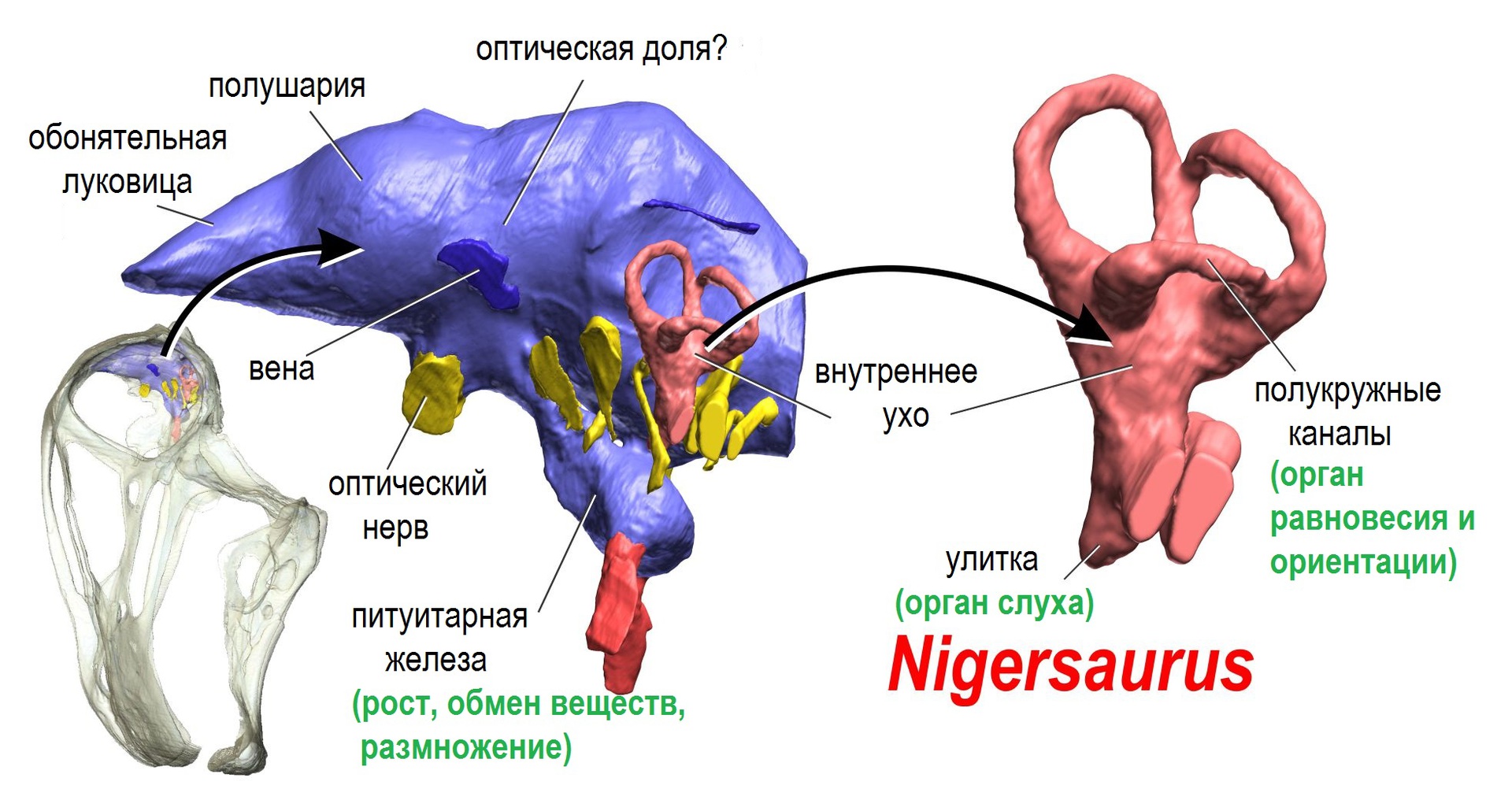
Центральная нервная система динозавровМозг тираннозавра считается одним из самых изученных для динозавров благодаря хорошей сохранности черепов данных животных. В целом структура его мозга аналогична современным крокодилам и имеет примерно схожий с взрослым аллигатором коэффициент энцефализации. Весьма высокое значение для таких крупных хищников своего времени, уступающее лишь некоторым прогрессивным дромеозаврам — бамбираптору, троодону и орнитомиму, чей КЭ был сравним со средним показателем современных птиц.Слепок черепной коробки тираннозавра и аллигатора. Обратите внимание на сильно развитую обонятельную луковицу обоих животных, вполне типичную для хищных пресмыкающихся (1). 2 — оптический нерв; 3 — большие полушария головного мозга; 4 — оптическая доля; 5 — мозжечок; 6 — вестибулярный аппарат; 7 — продолговатый мозг
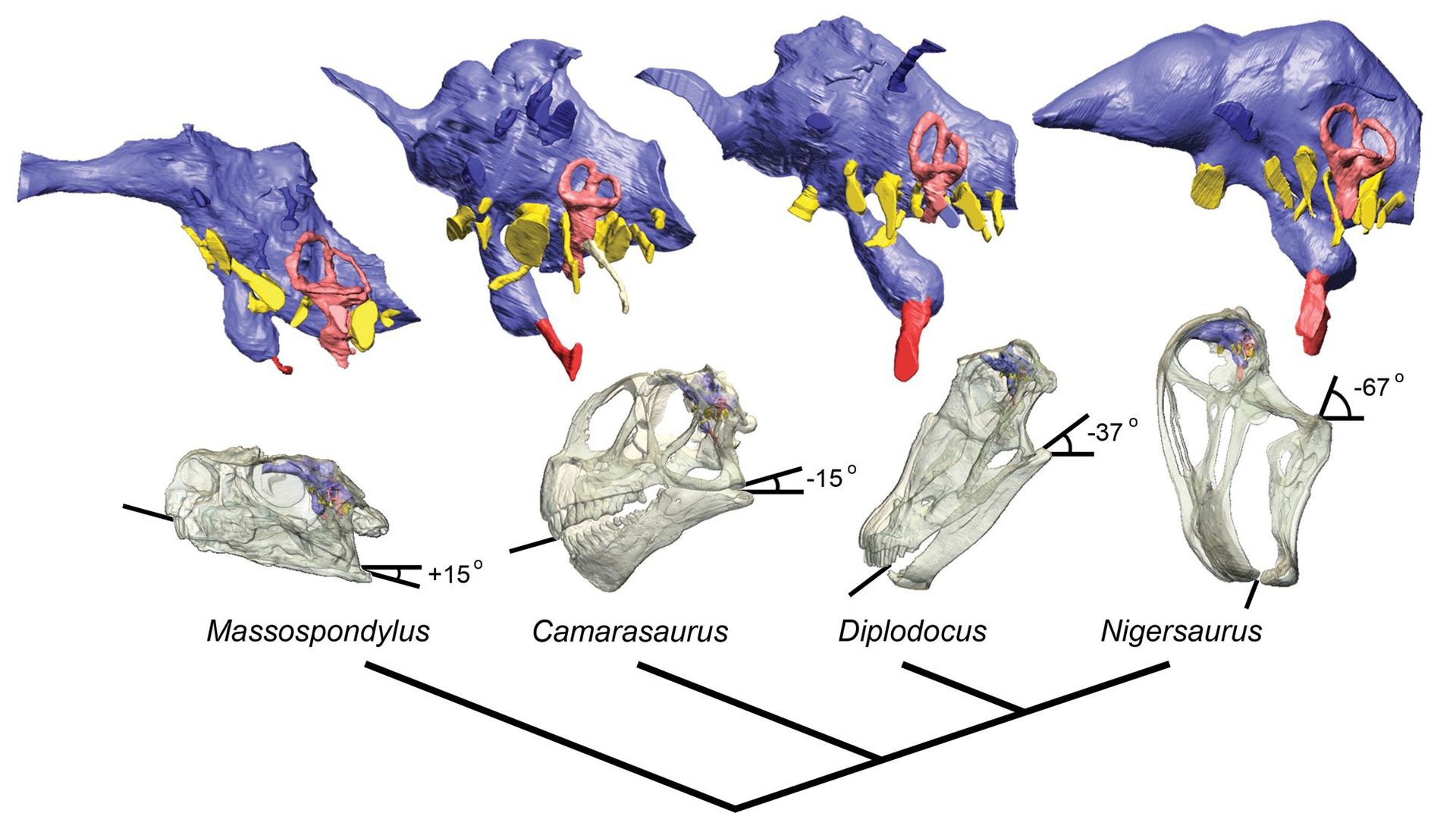
Слепки черепной коробки различных зауроподоморф (в градусах указанна растяжимость челюстного аппарата животного). Внизу описание структуры мозга Нигерозавра.
Мозг птиц и млекопитающих базируется на стриатуме, доставшийся им от пресмыкающихся предков.Тогда как млекопитающие пошли по пути увеличения объема и функциональности коры головного мозга (покрывающего стриатум), архозавры и позднее птицы пошли по обратному пути наращивания объемов и функциональности стриатума. Таким образом высшая нервная деятельность современных птиц обеспечивается стриатумом (точнее в гиперстриатуме, аналоге неокортекса у млекопитающих). Кора обеспечивает лишь второстепенные функции (предположительно обоняние). Аналогов гиперстратума у динозавров (пока?) не обнаружено, что опять таки позволяет предположить о менее развитом чем у высших птиц интеллекте теропод.
Эволюция мозга и обонятельных долей мозга от динозавров (бамбираптор) до современных птиц (голубя). Доли полушарий головного мозга птиц крупнее чем у динозавров, тогда как обонятельные доли существенно меньше. По мнению исследователей, это может быть связанно с высоким уровнем интеллекта птиц, компенсирующий отсутствие относительно сильного обоняния
График демонстрирующий коэффициент энцефализации для разных групп позвоночных. Коэффициент птиц отстоит от такового у своих предков-динозавров примерно на столько же, на сколько КЭ современных млекопитающих отстоит от значения коэффициента своих предков эпохи динозавров
«Кости судьбы»
Кости древних ящеров могут не только дать нам представление о внешнем виде динозавров (морфология), но так же «поведать» историю жизни отдельного животного. Речь, конечно же, не только в изучении повреждений костных тканей, которое животное пережило в течение своей жизни в результате травм и инфекций (как это описано в док. Фильме «Баллада о Большом Але», но так же в структуре самих костей.
Кости рептилий, подобно стволу деревьев, в срезе обнаруживают своеобразные «годовые кольца», отмечающие сезонные изменения в скорости роста животного. Лучше всего для таких целей обычно подходят плечевые кости или кости бедра динозавра. Подобный анализ (называемый гистологическим), позволяет оценить темпы роста животного, время достижения половой зрелости, пол и возраст на момент смерти.
Пример годовых колец на срезе костей травоядного Гиппакозавра. Внизу видны толстые светлые кольца ускоренного роста животного в первые годы жизни, разграниченные темными сезонными полосами (засухи или похолодание). Гистологический анализ оценивает достижение половой зрелости к 2–3 годам (при размерах в 40% от максимального для данного вида). Максимального размера (в 9 м длины и 4т веса) животное достигало к 10–12 годам. Такие темпы роста более характерны для современных птиц (страусов), нежели современных прогрессивных рептилий (крокодилов)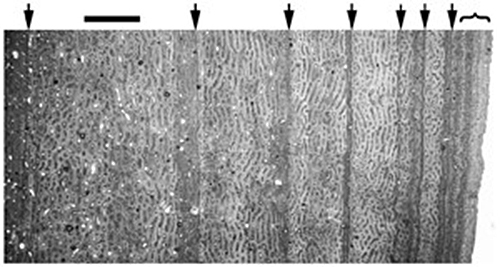
По следам медуллярной ткани в бедренной кости животного так же с высокой вероятностью возможно определение пола древнего ящера (она необходима самкам как резерв кальция, необходимого для скорлупы яиц). Подобная ткань откладывается в костях архозавров (у птиц она, как и у динозавров-теропод, запасается в бедренной кости) при достижении ими репродуктивного возраста. Половозрелость динозавра обычно коррелирует с резким замедлением роста животного. Так, медуллярная ткань была обнаружена в бедре самки тираннозавра из Монтаны (США) возрастом (при смерти) в 18 лет. Изучение других экземпляров тираннозавров (вида Рекс) так же фиксирует резкое замедление роста к 16–20 годам. Самому старому из обнаруженных экземпляров на момент смерти было около 28 лет. Факт того что большинство найденных останков тираннозавров рекс принадлежат особям прожившим лишь несколько лет после наступления половой зрелости, свидетельствует о весьма тяжелых условиях полового отбора этих созданий.
«Медуллярный принцип» позволяет делать и весьма интересные открытия. Один из обнаруженных овирапторов (сестринская по отношению к тираннорапторам группа целлурозавров) на момент смерти высиживал кладку подобно современным птицам. Гистологические исследования не обнаружили в костях животного медуллярной ткани (фото внизу справа), из чего сделали вывод что у данного вида кладку высиживал самец, или как минимум оба родителя
У многих видов современных птиц отцы тоже занимаются «воспитанием» своего потомства. Ниже снимок самца казуара со своими птенцами которых высиживал так же он сам. Животное напоминает овираптора даже внешне. На рисунке слева отчетливо виден длинный коготь на первом пальце ноги казуара, аналогично «ужасным когтям» дейнонихозавров. Однако в отличии от последних, используется он для защиты животного от хищников
«Экзамен» природы
Иногда природа преподносит нам подарок, позволяющий подкорректировать наши представления о динозаврах.
В 2000 году, в Северной Америке была найдена хорошо сохранившаяся «мумия» травоядного динозавра рода гадрозавров названная Дакотой. При анализе останков ученые выяснили что эти динозавры при жизни были на 1 м длиннее, а мышечная масса была на 25% больше чем считалось ранее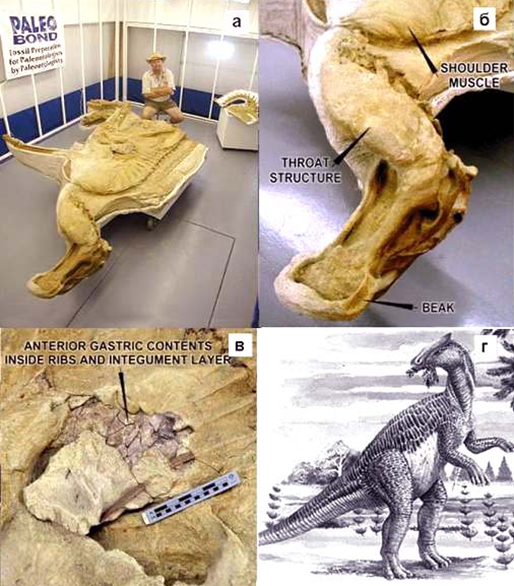
Касательно моделирования мышечной структуры динозавров, на видео можно ознакомиться на примере процесса виртуального «восстановления» мышц шеи и черепа аллозавра
Смерть встретила практически одновременно хищного велоцераптора и его жертву протоцератопса в момент борьбы, запечатлевшей поединок на 70 млн лет. Одна из ног хищника своим когтем находилась в области шеи протоцератопса, что дало дополнительный вес сторонникам теории о том что дейнонихозавры использовали большие когти на первых пальцах ног преимущественно для повреждения трахеи или артерии жертвы в шейном отделе, а не брюха или груди (как это делают казуары), для чего когти были не достаточно прочными
«Гены древних»
Да, вы верно поняли, костные останки динозавров так же могут дать нам представление об их геноме, точнее о размере их ДНК. Ученые, при изучении клеток костной ткани животных (остеоцитов), заметили линейную связь между размерами остеоцитов и ДНК животных. Это позволило им оценить и размеры геномов доисторических ящеров по сравнению с современными птицами и рептилиями. Так ДНК крокодила содержит чуть более 3 млрд. пар нуклеиновых оснований (НО), в то время как ДНК птиц в среднем содержит 1,45 млрд. пар НО (от 0,97 до 2,5 млрд). По оценкам ученых размеры ДНК динозавров сильно различаются у ящеротазовых и птицетазовых соответственно. Средний размер генома птицетазовых динозавров был близок к таковому у современных пресмыкающихся — 2,5 млрд. пар НО. Ящеротазовые же (тероподы и зауроподы) показывали значения более близкие к современным птицам 1.8 млрд. пар НО.
Размеры ДНК у разных типов живых организмов.
У позвоночных животных размер генома плохо коррелирует с числом генов. Ни размеры ДНК ни число генов не имеют положительной корреляции с уровнем развития живого существа. Так ДНК человека содержит около 3 млрд пар НО. кодирующие 25 — 30 000 генов, далеко не рекорд в животном мире. Подавляющая часть ДНК почти не участвует в жизнедеятельности клетки позвоночных (так называемый «генетический мусор»), и может быть существенно «урезанна» без серьезных последствий для животных.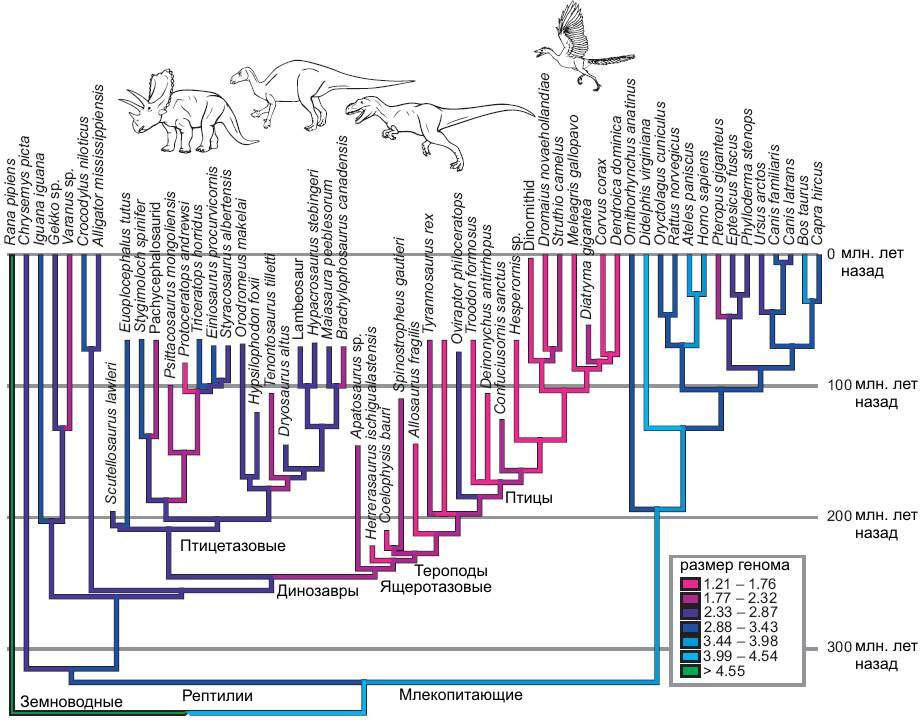
Выходит что птицетазовые и ящеротазовые динозавры, предположительно, пошли разными путями от общего триасового предка имевшего типично большой для рептилий геном. Ящеротазовые, в своем большинстве, освоили бипедальную локомоцию (тероподы), что потребовало радикального сокращения генома для оптимизации системы кровообращения (сокращение генома эритроцитов для уменьшения их размеров). Соответственно уже позднее, в юрском и меловом периоде такой геном сохранился у теропод-манирапторов, постепенно перешедших к активному полету — настоящих птиц.
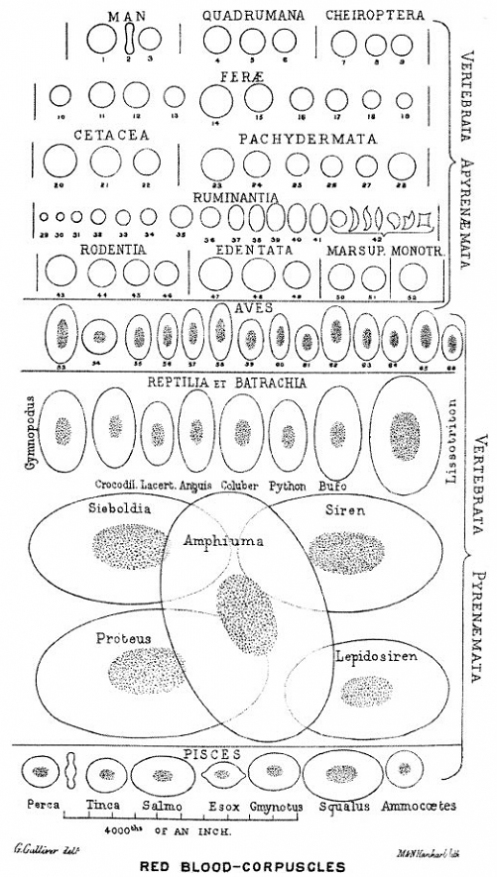
Эти данные являются дополнительными косвенными свидетельствами в пользу гипотезы появления современных пернатых от динозавров-теропод юрского периода. Впрочем, перья тоже не являются исключительно птичьей монополией, и наряду с костями древних ящеров оставили нам ископаемые свидетельства развитого перьевого «гардероба» которым располагали «истинные» динозавры.
В следующей части обзора речь пойдет об эволюции пернатого «гардероба» динозавров, их пути от простых украшений при брачных играх и до усвоения манирапторами полноценного полета.
Ссылки:
Ляонин, Провинция динозавров и электроники
Загадка «китайских Помпеев» решена
Кладистика
Какие жили динозавры на территории России
Переходные формы: как их нашли и не нашли
Грудная клетка и механизм дыхания у птиц
Эритроциты и размер генома
Малый размер птичьих геномов — наследие эпохи динозавров
«Эволюция. Классические идеи в свете новых открытий». Глава из книги
Evolution of the sense of smell in dinosaurs and birds
Чем «думают» птицы
Микрораптор — четырехкрылый динозавр
