Внеклеточная ДНК, как биомаркер старения и различных патологий
Мы уже рассказывали вам про циркулирующих микроРНК, но есть еще одна не менее крутая штука — внеклеточная циркулирующая ДНК (англ. cell-free DNA, circulating DNA). Открыта она была ещё 1948 году. Сейчас к ней усилился интерес и вот в чем собственно дело.
Внеклеточная ДНК (вкДНК) обнаруживается в нескольких жидкостях организма (в плазме и сыворотке крови, моче, слюне, синовиальной, перитонеальной и спинномозговой жидкости).
Мы с вами рассмотрим особенности вкДНК в двух жидкостях: крови и моче. Понятно дело, что так удобней для диагностики.
Источники вкДНК в организме.
Как стало известно, вкДНК циркулируют в крови в составе апоптотических телец, микровезикул, нуклеосом, экзосом, нуклеопротеиновых комплексов с белками крови и, предположительно, в свободном виде.
Считается, источниками возникновения вкДНК в крови являются процессы гибели клеток, созревание эритроцитов, секреция ДНК клетками, а также бактерии и вирусы. Сразу отметим, что последний источник, ДНК бактерий и вирусов, вносят очень незначительный вклад в общий пул циркулирующей ДНК, и уровень экзогенной ДНК во время инфекций не превышает нескольких десятков пкг/мл. Тогда как по современным данным, нормальный уровень вкДНК в здоровом организме составляет 3–44 нг/мл в плазме и 50–100 нг/мл в сыворотке крови.
Значительная часть вкДНК в организме вероятно появляется в результате гибели клеток, в процессе апоптоза, некроза и нетоза. Ежедневно в организме взрослого человека посредством программируемой клеточной гибели (апоптоза) гибнет около ста миллиардов клеток, в результате чего деградирует около 1 г. ДНК в сутки.
В процессе апоптоза ядерная ДНК расщепляется специальным ферментом ДНКазой. После чего появляются фрагменты ДНК длиной, кратной нуклеосоме, 180–200 п.н., которые в составе апоптотических телец (специальной упаковки после апоптоза) должны быть утилизированы макрофагами. П.Н. — пар нуклеотидов. Ну все знают, что пары нуклеотидов, аденин-тимин и гуанин-цитозин, в нашей ДНК повторяются три миллиарда раз. Нуклеосомы — специальные структуры в составе хроматина, состоящие из белков-гистонов. на которые в полтора оборота наматывается нить ДНК.
Дальше происходит странное. По непонятным причинам, часть ДНК погибших клеток избегает встречи с макрофагами и попадает в кровь. В разное время проводились эксперименты, подтверждавшие апоптотическую природу вкДНК [1,2].
При разделении вкДНК плазмы здоровых доноров с помощью гель-электрофореза обнаруживаются полосы, соответствующие длинам фрагментов 180–200 п.н. и длинам, кратным этому числу, что соответствует межнуклеосомному расщеплению хроматина при апоптозе.
Обнаружение ДНК плода в крови матери также говорит в пользу апоптоза как источника вкДНК [3].
В норме некроз вносит менее значительный вклад в появление вкДНК, увеличивая её количество при тяжёлых травмах. Некротическая вкДНК в плазме обнаруживается в виде более длинных фрагментов — длиной свыше 10 тысяч п.н. С помощью метода ПЦР (полимеразной цепной реакции) возможно различить длинные фрагменты вкДНК, появляющиеся в результате некроза и более короткие фрагменты — из-за апоптоза.
Кроме апоптоза и некроза существует ещё один малоизвестный вид клеточной гибели, являющийся источником вкДНК — нетоз (NETosis).
При нетозе клетки нейтрофилы выбрасывают специальные сетеподобные структуры, внеклеточные нейтрофильные ловушки (NET, Neutrophil Extracellular Trap), задача которых — дезактивация проникшего в организм патогена: вирусов, грибов и бактерий. При этом нейтрофил гибнет, выбрасывая ДНК, гистоны, различные белки и ферменты во внеклеточное пространство. Известно, что иногда процесс NET носит патологический характер, способствуя развитию тромбоза, сердечно-сосудистых, аутоиммунных заболеваний, а также рака.
Недавно, в 2018 году было обнаружено, что при ВИЧ-инфекции происходит гиперактивация нейтрофилов, механизма NET и нетоза. В результате чего происходила обширная гибель иммунных клеток, захваченных сетями нейтрофилов, CD4+ и CD8 + Т-клеток, В-клеток и моноцитов, и развитие сопутствующих сердечно-сосудистых патологий [4].
Ещё один потенциальный источник вкДНК — секреция ДНК нормальными и опухолевыми клетками. Ещё в 1972 году было получено доказательство активного выделения ДНК лимфоцитами во внеклеточную среду [5].
Наличие такое феномена, как выделение ДНК клетками во внеклеточную среду предполагает существование специальных механизмов переноса ДНК через клеточную мембрану. В ходе исследований было установлено, что высвобождение ДНК из лимфоцитов происходит при активном действии трипсина, проназы и плазмина на фоне дефицита ионов Са2+ и Mg2+. Избыток же ионов кальция ингибирует выход ДНК из клетки [6].
По мнению российских биохимиков из Казанского государственного университета, ещё одним вероятным источником вкДНК в крови может быть совсем неизвестная форма цитоплазматической ДНК, отличная от ядерной и митохондриальной — ДНК, ассоциированная с мембраной диплоидных лимфоцитов человека (Membrane-Associated DNA in the Cytoplasm of Diploid Human Lymphocytes) [7].
Несмотря на то, что этот тип ДНК был открыт в 1971 году, он и сейчас остаётся неизученным, и по нему есть считанные работы одних и тех же авторов, хотя и в PNAS и Nature.
По предположению Абрамовой З.И. с соавторами какая-то часть вкДНК может формироваться из этой цитоплазматической ДНК, на что указывают характерные особенности некоторых фрагментов вкДНК, отличные от ядерной и митохондриальной [6].
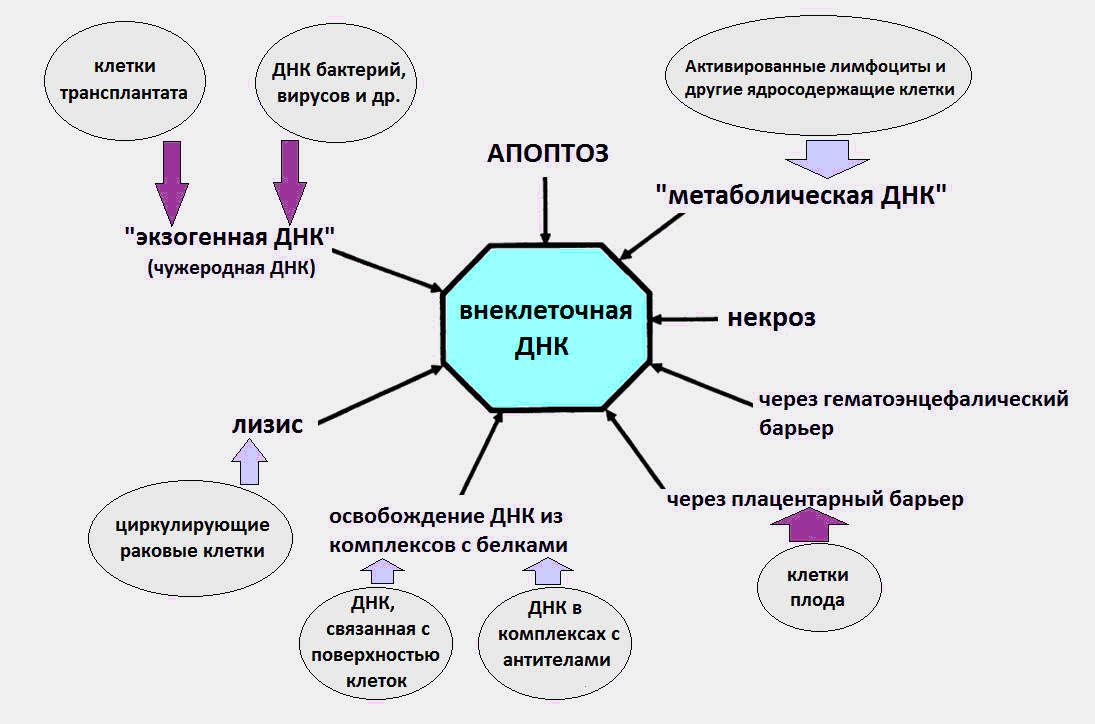
Рис. 1 Различные пути, которыми вкДНК попадает в циркуляцию.
Более подробно обо всех аспектах биологии вкДНК (иммуномодулирующих эффектах, рецепции к вкДНК на поверхности клеток и пр.) можно прочесть в подробном обзоре профессора Козлова В.А. (НИИ клинической иммунологии СО РАМН) [8].
Внеклеточная ДНК как биомаркер при различных патологиях
Логика подсказывает, что уровни вкДНК должны изменяться при патологиях, которые сопровождает массовая клеточная гибель. В ряде исследований было показано значимое повышение уровней вкДНК при развитии аутоиммунных заболеваний (системной красной волчанки, системной склеродермии, ревматоидного артрита и др.), и что существуют характерные особенности вкДНК, типичные для определённой аутоиммунной патологии [8–10].
Также известно, что при остром инфаркте миокарда, который сопровождает гибель клеток сердца по типу как некроза, так и апоптоза, уровни вкДНК также повышаются. И это имело потенциальное диагностическое значение, поскольку у пациентов, у которых развились осложнения, были более высокие уровни, чем у тех, кто восстанавливался лучше [11].
У больных, перенёсших острый инсульт, концентрация вкДНК в плазме крови, измеренная в течении 24 часов, коррелирует с тяжестью инсульта и может служить предиктором смертности и тяжести исхода даже у тех пациентов, у которых нет видимых изменений, обнаруживаемых методами нейровизуализации [12,13].
Известно, что для вкДНК характерно наличие большое количества гуанин-цитозин (ГЦ)-последовательностей, которые более устойчивы к разрушению нуклеазами, чем аденин-тимин-фрагменты ДНК. Так, содержание специфической ГЦ-обогащенной последовательности генома — транскрибируемой области рибосомного повтора — в составе вкДНК в несколько раз превышает содержание этого повтора в ядерной ДНК [14].
А при хронических патологиях, сопровождающихся активацией клеточной гибели (ишемической болезни сердца, артериальной гипертонии, аутоиммунных патологиях), содержание этих ГЦ-богатых маркерных последовательностей в вкДНК крови увеличивается в 10 и больше раз [15].
Что даёт полную возможность использовать показатели ГЦ-обогащённой рибосомной ДНК в составе вкДНК в качестве биомаркера хронически протекающего патологического процесса.
При борьбе с раком стоит задача обнаружить опухоль на самой ранней стадии, когда её только вообще можно обнаружить.
Тут вкДНК представляет крайний интерес, как биомаркер опухолегенеза.
Точечные мутации в опухолевой вкДНК обнаруживались в крови пациентов задолго до постановки диагноза[16].
Про уровни вкДНК при онкологии, поиску в составе вкДНК характерных для канцерогенеза мутаций, таких, как онкогены KRAS, TP53, HER2, BRCA1, APC, PIK3CA, BRAF и др., использованию в качестве маркера митохондриальной ДНК в составе вкДНК, изучению характерных генетических и эпигенетических изменений вкДНК, таких, как гипометилирование подвижных элементов Alu, при возникновении опухолей, использования «индекса целостности» вкДНК и прочих аспектов можно прочесть в работах Васильевой и соавт., Gonzalez-Masia и соавт. [17, 18].
Установлено, что для вкДНК характерно гораздо более сильная степень окисления под воздействием активных форм кислорода (АФК), чем для ядерной ДНК.
Так, содержание хорошо известного маркера окисления ДНК, 8-охоdG, в составе ядерной ДНК в норме и при различных патологиях колеблется от 1 до 10 на миллион нуклеотидов, а в составе вкДНК содержание этого маркера составляет 300 и более на миллион нуклеотидов [19].
Резкое повышение содержание 8-охоdG во вкДНК установлено при онкологии и сердечно-сосудистых заболеваниях, которые сопровождаются окислительным стрессом, и может достигать значений в 3000 8-oxodG на миллион нуклеотидов [20].
В целом, считается, что уровни 8-охоdG в составе вкДНК представляют собой чувствительный маркер общего окислительного стресса в организме.
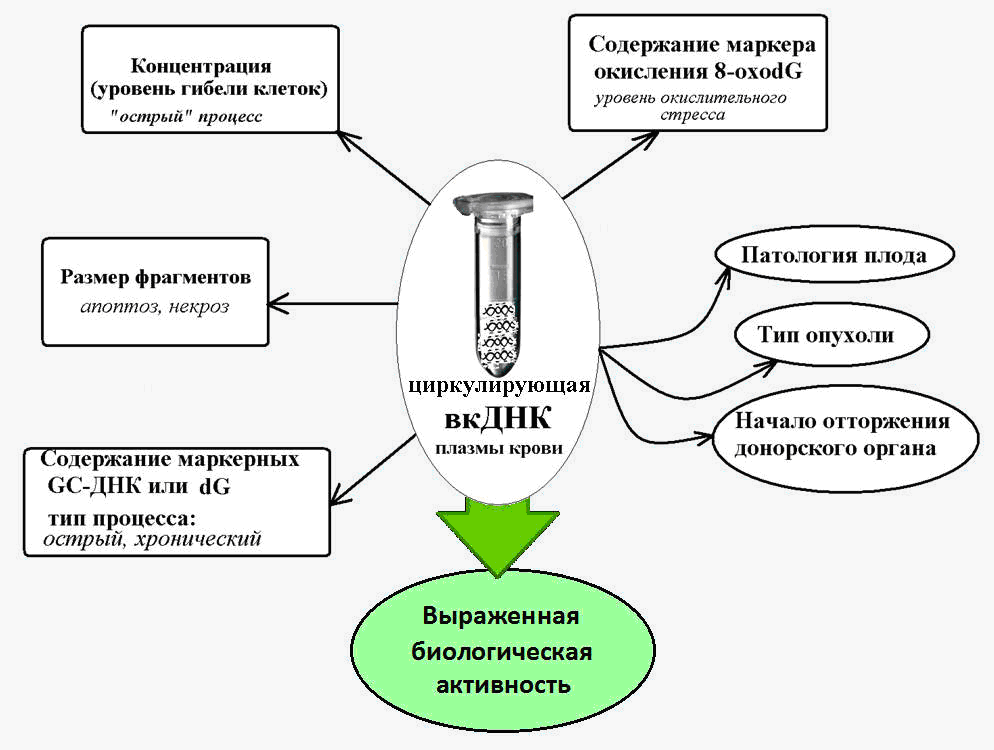
Рис. 2. Циркулирующая внеклеточная ДНК — маркер при различных патологических состояниях, сопровождающихся повышенной гибелью клеток.
Внеклеточная ДНК как мобильный элемент.
Большой и очень важный прорыв в исследованиях роли вкДНК во внутренних процессах произошёл 3 года назад. В 2015 году группой исследователей было сделано захватывающее открытие, показавшее, что вкДНК, подобно мобильным генетическим элементам, способна внедряться в ядерную ДНК и вызывать мутации в геноме [21]. Всё это очень удивительно.
В более ранних работах уже было показано, что опухолевая ДНК из крови может иметь способность встраиваться в ядерную ДНК. Мышиные фибробласты помещали в плазму, взятую у пациентов с раком толстой кишки, после чего в ДНК клеток мыши обнаруживались онкогены K-ras и гены белка р53, участвующего в апоптозе, а сами клетки были онкогенно трансформированы (при введении здоровым мышам вызывали рак) [22].
Что свидетельствует о том, что ДНК из плазмы была поглощена и интегрирована помещёнными в неё клетками. Это предполагаемое явление, поглощение здоровыми клетками онкогенов из циркулирующей в крови вкДНК, может быть ответственно за процесс метастазирования и получило название «генометастаз».
В другой работе были предоставлены данные о поглощении и интеграции фрагментов гена DYS14, специфичного для Y-хромосомы, в мозге женщины, вынашивавшей плод мужского пола [23].
Итак, в исследовании 2015 года учёные проследили, как флуоресцентно меченые вкДНК, взятые у больных раком и здоровых людей, без проблем встраивались в ядерную ДНК мышиных фибробластов в опытах in vitro (т.е вне живого организма, в клеточных культурах).
Причём человеческая вкДНК локализовалась в ядрах мышиных клеток в течение нескольких минут, и почти 100% ядер клеток мыши показывали флуоресцентные сигналы в течение 30 минут.
Секвенирование всего генома мышиных клеток показало присутствие в нём фрагментов ДНК человека, включая характерные мобильные элементы Alu. Кроме этого, авторы обнаружили индукцию γ-H2AX-очагов, возникающих при повреждении ДНК, а также повышение активности каспазы-3 и экспрессии генов ATM, p53, p21, GADD-34 и ДНК-лигазы. Что всё вместе свидетельствует о повышенной нестабильности генома, активации реакции ответа на повреждения ДНК и апоптоза.
Затем авторы проверили действие чужеродной вкДНК in vivo, на живом организме. Мышам внутривенно вводили человеческую вкДНК, фрагменты которой затем обнаружили в сердце, легких, печени и мозге мышей, умерщвленных через 7 дней после инъекции.
Причём ведённые мышам частицы человеческой ДНК оказались способны преодолевать гематоэнцефалический барьер и проникать в мозг. Как и в случае с клетками в культуре, внутривенное введение вкДНК вызывало индукцию γ-H2AX, проапоптотической каспазы-3 и активацию ответа на повреждения ДНК. И именно ответ на повреждение ДНК, по мнению учёных, играет центральную роль интеграции вкДНК в ядерную.
Ядерная ДНК воспринимает внедрение внеклеточной как двухцепочечный разрыв и начинает «латание» самой себя, встраивая циркулирующую ДНК в свой состав: «Активирование ответа на повреждение ДНК объединяет несколько разнородных вкДНК и фрагментированный хроматин в длинные конкатемеры посредством негомологичного соединения концов в рамках процесса восстановления».
В итоге авторы приходят к парадоксальному выводу: «Будучи повсеместным и постоянно возникающим, повреждение ядерной ДНК, вызванное внеклеточной циркулирующей ДНК, может быть основной причиной старения, связанных со старением отклонений и окончательной гибели организма. Таким образом, ДНК, возможно, играет парадоксальную роль как хранителя основ жизни, так и разрушителя самой жизни».
Надо ли говорить, что этот вывод очень сильно противоречит всем представлениям о старении на сегодняшний день.
Логика подсказывает, что этой гипотезе соответствовала бы следующая зависимость — чем больше продолжительность жизни особей вида, тем меньше у них этой формы ДНК и/или она для них менее опасна.
Но до тех пор, пока такое сравнительное исследование не выполнено, остается полагать, что у долгоживущего вида (человек) и короткоживущего (мышь) вкДНК производятся, предположительно, в относительно сравнимых уровнях. И обнаружение повышенных концентраций этих молекул у пожилых вполне может быть наоборот следствием процесса старения, а не его причиной.
Хотя это такое следствие, которое может усиливать причину. Читай статью Беликова.
Кроме этого, авторы открытия предлагают отнести вкДНК к ещё одному классу мобильных генетических элементов. И тогда возможно получит своё объяснение широкое участие в эволюции такого процесса, как горизонтальный перенос генов, как у беспозвоночных, так и позвоночных представителей животного мира. Благодаря которому, по одним из последним данным, геном человека мог заимствовать у других видов несколько десятков генов [24]
Остаётся открытым очень важный вопрос: как именно вкДНК попадают в клетку? Имеющиеся гипотезы включают предположения, что частицы, содержащие ДНК, могут проникать в клетки через имеющиеся в мембране поры, а также посредством разных типов эндоцитоза, протеогликан-зависимого макропиноцитоза или при взаимодействии с мембранными белками.
Внеклеточная ДНК при ожирении, воспалении, рентгеновском облучении и стрессе различной природы
Ещё одно интересное открытие было сделано год спустя японскими исследователями. Они смогли описать активное участие вкДНК, появившейся в крови в результате гибели адипоцитов у людей с ожирением, в развитии воспаления и резистентности к инсулину. Ключевым фактором, взаимодействующим в этих процессах с вкДНК, была молекула врождённого иммунитета Toll-подобный рецептор 9 (TLR9) [25].
Не обошлось и без интриги со стороны российских учёных из Института теоретической и экспериментальной биофизики РАН. Они установили, что рентгеновское облучение и антидиабетический препарат метформин вызывают увеличение выведения с мочой вкДНК (ядерной и митохондриальной) у крыс.
Их данные показали, что до опытов количество ядерной вкДНК было на 40% выше, а митохондриальной вкДНК — на 50% выше в моче пожилых крыс по сравнению с молодыми.
Через 12 ч после облучения содержание я-вкДНК и мт-вкДНК в моче молодых крыс увеличилось на 200% и 460% соответственно по сравнению с контролем, тогда как в моче пожилых крыс оно стало на 250%. и 720% выше.
Через 6 ч после введения метформина другим крысам, количество я-вкДНК и мт-вкДНК в моче молодых крыс было повышено на 25% и 55% и на 50% и 160% в моче пожилых крыс.
По мнению авторов, эти данные говорят о том, что рентгеновские лучи и метформин вызывают значительное увеличение вкДНК в моче крыс (особенно старых), вызванное активной гибелью клеток в тканях. Эти результаты также позволяют предположить, что метформин, возможно, действует как сенолитик, инициируя гибель старых клеток, содержащих структурные и функциональные нарушения [26].
В октябре прошлого года, учёные проследили взаимосвязь уровней вкДНК с психическим стрессом и физическими нагрузками у молодых людей в возрасте 18–36 лет. Результаты показали двукратное увеличение вкДНК после острого психосоциального лабораторного стресса (TSST) и пятикратное увеличение вкДНК после упражнений на беговой дорожке (с достижением истощения сил бегущего).
Было также зафиксировано повышенное количество более коротких фрагментов вкДНК, характерное для апоптоза, в крови после физического истощения.
Внеклеточная митохондриальная ДНК показала одинаковое увеличение после обеих стрессовых нагрузок. Кроме этого, профили метилирования вкДНК, используемые в этой работе в качестве маркера для различного клеточного происхождения, значительно различались для разных стресс-тестов.
Для анализа метилирования учёными был выбран активно участвующий в эмбриональном развитии ген HOXA5 из-за специфичности его метилирования в разных типах клеток. Так, для него характерно гиперметилирование в мышечных клетках, гипометилирование в клетках мозга, из клеточных линий гиппокампа и культивируемых нейронов, и дифференциальное метилирование в клетках крови.
Метилирование гена HOXA5 во вкДНК понижалось сразу после психосоциального стресса и увеличивалось после физического стресса, что, по мнению авторов, указывает на различные клеточные источники активного высвобождения ядерной ДНК. [27].
Внеклеточная ДНК и старение.
Теперь главное. Рассмотрим взаимосвязь уровней и специфических особенностей вкДНК со старением.
Первые работы, показавшие такую взаимосвязь, были опубликованы в 2011–13 годах. Финские микробиологи и иммунологи из университета Тампере изучили показатели вкДНК у долгожителей старше 90 лет, участников исследования Vitality 90+, и у молодых людей (в возрасте 22–37 лет) в качестве контрольной группы.
Результаты их работ показали, что концентрация вкДНК у долгожителей была значительно выше, чем у молодых. Также были характерные различия вкДНК: у старых людей она была представлена больше низкомолекулярными фрагментами, у молодых — больше высокомолекулярными, то есть более длинными.
В следующих работах эта же группа показала, что уровни вкДНК имели чёткую взаимосвязь с маркерами воспаления (положительно коррелировали с уровнем С-реактивного белка, сывороточного амилоида А (SAA) и др.) и смертностью у людей старше 90 лет.
А также то, что более высокие уровни общей и гипометилированной вкДНК были связаны с системным воспалением и развитием старческой астении (frailty) [28–30].
Два самых свежих последних прошлогодних исследования по вкДНК и старению показали характерные эпигенетические изменения этой ДНК.
В одном из них команда учёных проанализировала, как с возрастом изменяется метилирование мобильных генетических элементов LINE-1 и Alu во вкДНК здоровых людей, в возрасте от 23 до 61 года. Элемент LINE-1 имеет длину около 6 тыс.п.н., тогда как высокомолекулярные фрагменты вкДНК, для сравнения, имеют размер более 10 тыс. п.н.
LINE-1 относится к ретроэлементам, т.е. к мобильным элементам, которые для своего размножения используют обратную транскрипцию, перенос генетической информации с РНК на ДНК. LINE-1 — единственный известный на сегодня мобильный элемент генома, который сохраняет способность к размножению и перемещению своих копий в геноме. Также это самый многочисленный из подвижных структур человека: копии LINE-1 занимают в ДНК огромную долю — пятую часть всего генома.
Второй элемент, Alu, не может самостоятельно размножаться и делать новые вставки в геном, и зависит в этом процессе от генетического аппарата элемента LINE-1 (подробнее о ретроэлементах генома см. статью в Популярной Механике «Ретровирусы: «пятая колонна» ДНК» www.popmech.ru/science/178711-retrovirusy-pyataya-kolonna-dnk/#part0)
Результаты исследования показали, что по мере старения происходит значимое уменьшение метилирования ретроэлементов LINE-1 и Alu, причём этот процесс заметно интенсивнее проходил у курильщиков.
Как сегодня установлено, активность элементов LINE-1 и Alu, которые способны вставлять свои копии в геном, вносят вклад в значительную часть генетических заболеваний человека и рака.
Известно о более 100 опосредованных LINE-1 инсерциях (вставках), которые приводят к генетическим заболеваниям человека и найдено несколько вставок LINE-1, которые прерывают гены-супрессоры опухолей и провоцируют рак [31].
Во втором исследовании также учёные подтвердили снижение с возрастом метилирования элементов LINE-1 и Alu в составе вкДНК. Причём, среди 4 групп в исследовании (здоровые долгожители старше 100, нездоровые долгожители старше 100 лет, пожилые люди, 71 год и молодые люди, 25 лет) схожие характеристики вкДНК имели здоровые долгожители с молодыми людьми и пожилые люди с нездоровыми долгожителями:
«Постоянно в нашем исследовании мы отмечали большее сходство в профилях вкДНК, как глобально, так и локально между молодыми и здоровыми долгожителями, в отличие от пожилых людей и нездоровых долгожителей. Следовательно, наше исследование предполагает, что профилирование вкДНК может быть использовано не только в качестве биомаркера возраста, но и в качестве предиктора общего состояния здоровья». [32].
Таким образом, как мы видим, внеклеточная циркулирующая ДНК оказалась тесно связана с состоянием здоровья и процессами, происходящими в организме в ходе старения: окислительным стрессом, воспалением и возрастзависимыми патологиями.
Современные методы исследования, такие, как количественная ПЦР в реальном времени, позволяют использовать вкДНК в качестве очень чувствительного биомаркера процессов, связанных со старением.
Проведённые исследования показали большую потенциальную ценность анализа вкДНК в диагностике рака, в траспланталогии, при сердечно-сосудистых заболеваниях и болезнях почек, фиброзе, в пренатальной диагностике (во время беременности), при травмах и сепсисе, а также в спортивной медицине.
Полученные данные о том, что вкДНК может проявлять свойства мобильного элемента и встраиваться в ядерную ДНК, тем самым влияя на развитие связанных с мутациями патологий и старением в целом, также представляют несомненный интерес.
Всё это требует дальнейших исследований и уточнений.
Обзор подготовили: Алексей Ржешевский и немного Михаил Батин
- Jahr S, Hentze H, Englisch S, Hardt D, Fackelmayer FO, Hesch RD, Knippers R. DNA fragments in the blood plasma of cancer patients: quantitations and evidence for their origin from apoptotic and necrotic cells. Cancer Res. 2001 Feb 15;61(4):1659–65.
- Deligezer U1, Yaman F, Erten N, Dalay N. Frequent copresence of methylated DNA and fragmented nucleosomal DNA in plasma of lymphoma patients. Clin Chim Acta. 2003 Sep;335(1–2):89–94.
- Bishoff F.Z., Dang D., Horne C., Marquez-Do D., Brincley W.R., Levis D.E. Fetal DNA in maternal plasma circulates as apoptotic bodies: elucidations of structural nature of fetal DNA for non-invasive prenatal genetic diagnosis // Am. J. Hum. Genet. 2003. V. 73. P. 189.
- Sivanandham R, Brocca-Cofano E, Krampe N, Falwell E, Venkatraman SMK, Ribeiro RM, Apetrei C, Pandrea. Neutrophil extracellular trap production contributes to pathogenesis in SIV-infected nonhuman primates. J Clin Invest. 2018 Nov 1;128(11):5178–5183.
- Rogers J.C., Boldt D., Kornfeld S. Excretion of deoxyribonuclein acid by lymphocytes stimulated with phytohemagglutinin or antigen // Proc. Nat. Acad. Sci. USA. 1972. V. 69, No 7. P. 1685–1689.
- Туаева Н.О, Абрамова З.И. Внеклеточная ДНК в кровотоке человека. 2007. Учебные записки Казанского государственного университета. Т.149, с. 23–32.
- Lerner R.A., Meinke W., Goldstein D.A. Membrane-associated DNA in the cytoplasm of diploid human lymphocytes // Proc. Nat. Acad. Sci. USA. 1971. V. 68, No 6. P. 1212–1216.
- Козлов В.А. Свободная внеклеточная ДНК в норме и при патологии. Медицинская иммунология. 2013;15(5):399–412.
- Galeazzi M, Morozzi G, Piccini M, Chen J, Bellisai F, Fineschi S, Marcolongo R. Dosage and characterization of circulating DNA: present usage and possible applications in systemic autoimmune disorders. Autoimmun Rev 2003;2:50–5.
- Leon SA, Revach M, Ehrlich GE, Adler R, Petersen V, Shapiro B. DNA in synovial fluid and the circulation of patients with arthritis. Arthritis Rheum 1981;24:1142–50.
- Destouni A, Vrettou C, Antonatos D, Chouliaras G, Traeger-Synodinos J, Patsilinakos S, et al. Cell-free DNA levels in acute myocardial infarction patients during hospitalization. Acta Cardiol 2009;64:51–7.
- Rainer TH, Wong LKS, Lam W, Yuen E, Lam NYL, Metreweli C, Lo YMD. Prognostic Use of Circulating Plasma Nucleic Acid Concentrations in Patients with Acute Stroke. Clin Chem 2003; 49:562–9.
- Lam NY, Rainer TH, Wong LK, Lam W, Lo YM. Plasma DNA as a prognostic marker for stroke patients with negative neuroimaging within the first 24 h of symptom onset. Resuscitation 2006; 68:71–8.
- Вейко Н.Н., Булычева Н.В., Рогинко О.А., Вейко Р.В., Ершова Е.С., Коздоба О.А., Кузьмин В.А., Виноградов А.М., Юдин А.А., Сперанский А.И., Фрагменты транскрибируемой области рибосомного повтора в составе внеклеточной ДНК — маркер гибели клеток организма, Биомедицинская химия, 2008, том: 54(1), 78–93.
- Bulicheva, N. et al. Effect of cell-free DNA of patients with cardiomyopathy and rDNA on the frequency of contraction of electrically paced neonatal rat ventricular myocytes in culture. Ann N Y Acad Sci. 2008. V. 1137. P. 273–277.
- Lecomte, T. Circulating free tumor DNA and colorectal cancer/ T. Lecomte, N. Ceze, E.
- Dorval, P. Laurent-Puig // Gastroenterol Clin Biol. 2010. V. 34. № 12. P. 662- 681.
- Васильева И.Н, Беспалов В.Г. Роль внеклеточной ДНК в возникновении и развитии злокачественных опухолей и возможности её использования в диагностике. Вопросы онкологии. 2013.-N 6.-С. 673–681
- Gonzalez-Masia, J.A. et al. Circulating nucleic acids in plasma and serum (CNAPS): applications in oncology. Onco Targets Ther. 2013. V. 8. № 6. P. 819–32.
- Ermakov, A.V. et al. Oxidized extracellular DNA as a stress signal in human cells. Oxid Med Cell Longev. 2013. V. 2013. P. 649–747.
- Loseva, P. et al. Extracellular DNA oxidation stimulates activation of NRF2 and reduces the
- production of ROS in human mesenchymal stem cells. Expert Opin. Biol. Th. 2012. V. 12. № 1. P. 85–97.
- Mittra I, Khare NK, Raghuram GV, et al. Circulating nucleic acids damage DNA of healthy cells by integrating into their genomes. J Biosci. 2015;40(1):91–111.
- Garcia‐Olmo DC, Dominguez C, Garcia‐Arranz M, Anker P, Stroun M, Garcia‐Verdugo JM, Garcia‐Olmo D (2010) Cell‐free nucleic acids circulating in the plasma of colorectal cancer patients induce the oncogenic transformation of susceptible cultured cells. Cancer Res. 70, 560–567
- Chan W, Gurnot C, Montine TJ, Sonnen JA, Guthrie KA, Lee Nelson J (2012) Male Microchimerism in the Human Female Brain. PLoS ONE 7, e45592.
- Crisp A, Boschetti C, Perry M, et al.: Expression of multiple horizontally acquired genes is a hallmark of both vertebrate and invertebrate genomes. Genome Biol. 2015;16:50. 10.1186/s13059–015–0607–3
- Sachiko Nishimoto, Daiju Fukuda et al. Obesity-induced DNA released from adipocytes stimulates chronic adipose tissue inflammation and insulin resistance. Sci Adv. 2016 Mar; 2(3): e1501332.
- Gaziev A, Abdullaev S, Minkabirova G, Kamenskikh K. X-rays and metformin cause increased urinary excretion of cell-free nuclear and mitochondrial DNA in aged rats. J Circ Biomark. 2016 Oct 25; 5:1849454416670782.
- Hummel EM, Hessas E, Müller S, Beiter T, Fisch M, Eibl A, Wolf OT, Giebel B, Platen P, Kumsta R, Moser DA Transl Psychiatry. 2018 Oct 29;8(1):236.
- Jylhävä J1, Kotipelto T, Raitala A, Jylhä M, Hervonen A, Hurme M. Aging is associated with quantitative and qualitative changes in circulating cell-free DNA: the Vitality 90+ study.
- Mech Ageing Dev. 2011 Jan-Feb;132(1–2):20–6.
- Jylhävä J1, Jylhä M, Lehtimäki T, Hervonen A, Hurme M. Circulating cell-free DNA is associated with mortality and inflammatory markers in nonagenarians: the Vitality 90+ Study. Exp Gerontol. 2012 May;47(5):372–8.
- Jylhävä J1, Nevalainen T, Marttila S, Jylhä M, Hervonen A, Hurme M. Characterization of the role of distinct plasma cell-free DNA species in age-associated inflammation and frailty. Aging Cell. 2013 Jun;12(3):388–97.
- Lars Erichsen, a Agnes Beermann et al. Genome-wide hypomethylation of LINE-1 and Alu retroelements in cell-free DNA of blood is an epigenetic biomarker of human aging. Saudi J Biol Sci. 2018 Sep; 25(6): 1220–1226.
- Teo YV, Capri M, Morsiani C, Pizza G, Faria AMC, Franceschi C, Neretti N. Cell-free DNA as a biomarker of aging. Aging Cell. 2019 Feb;18(1): e12890.
