Липкие гекконы: адгезионный липидный слой

Достаточно беглого взгляда на некоторых представителей флоры и фауны, чтобы убедиться в безграничности возможных исходов эволюции. Физиологические особенности тех или иных существ могут казаться нам странными, но для их обладателей — это в первую очередь функциональные атрибуты, необходимые для выживания. Не удивительно, что многие природные умения различных существ стали объектом научных исследований в области бионики, когда человек пытается воспроизвести их для дальнейшего применения в создании новых материалов и устройств. Летучие мыши, пауки, медузы и многие-многие другие обитатели Земли стали источником вдохновения в бионике благодаря своим удивительным особенностям. И гекконы не исключение. Ученые из Национального института стандартов и технологий (США) изучили умение гекконов прилипать к любым поверхностям, установив, что секрет этого таланта не только в необычной структуре поверхности их лапок, но и в специальных молекулах. Что это за молекулы, как ученые их нашли, и насколько они важны для гекконов, чтобы не падать с потолка? Ответы на эти вопросы мы найдем в докладе ученых. Поехали.
Основа исследования
Гекконы на самом деле это не вид, а очень многочисленное семейство, насчитывающее порядка 950 видов. Обитают они преимущественно в теплых регионах планеты. Рацион гекконов по большей степени состоит из насекомых и сока плодов, но есть виды, которые не брезгуют перекусить своими более мелкими сородичами.

Геккон вида токи (Gekko gecko).
Гекконы обладают удивительной способностью ползать по любым поверхностям в любых условиях. Они могут взобраться по стене и бегать вниз головой по потолку, как будто для них законы физики не писаны. Этот талант используется для побега от хищников, ловли добычи (обычно насекомых) и для очень драматичных боев за территорию или потенциального партнера, напоминающих ту самую сцену из фильма »300 спартанцев».

Тут отчетливо видны те самые «липкие» подушечки на пальцах геккона.
На подушечках пальцев гекконов имеются микроскопические волоски (щетинки), которые и обеспечивают им сцепление с поверхностью (изображение №1).
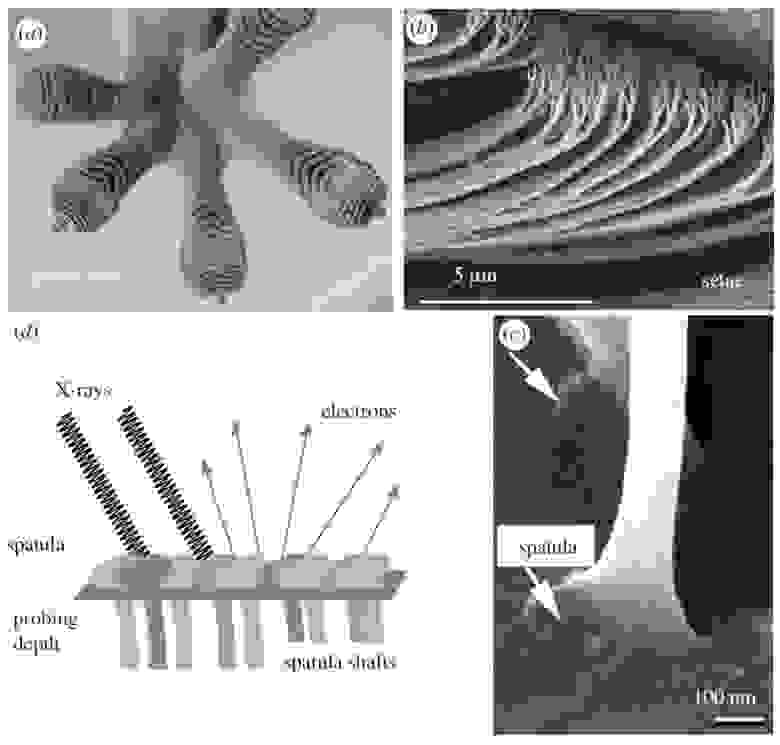
Изображение №1
Каждая щетинка представляет собой пучок спатул с уплощением на конце, что обеспечивает адгезивный контакт с поверхностью. Ветви щетинок происходят из наружного эпидермального слоя чешуи геккона. Массивы щетинок в основном состоят из β-белков, которые обеспечивают жесткость ткани, необходимую для постоянного прикрепления и отсоединения.
Механизм адгезии гекконов был в центре внимания исследований в области биомеханики и материаловедения в течение многих лет. Некоторые исследования пришли к выводу, что сцепление щетинок геккона к поверхности обусловлено взаимодействием Вандер-Ваальса, в то время как другие труды отметили заметную роль воды. В этих и других ранних исследованиях игнорировалась химия поверхности щетинок, так как считалось, что она имеет второстепенное значение.
Сейчас же можно уверенно сказать, что химия все же важна. К примеру, вибрационная спектроскопия и ядерный магнитный резонанс указывают на важную роль липидов и кислотно-основных взаимодействий в адгезии гекконов.
Ученые отмечают, как и в любом явлении адгезии, внешняя молекулярная структура щетинок определяет механический и химический контакт во время адгезии. Следовательно, подробная информация о химическом состоянии поверхности щетинок может быть крайне полезна для будущих бионических применений.
Недавно было замечено, что массивы щетинок геккона имеют связанные с ними липиды. Известно, что липиды располагаются в эпидермисе рептилий по схеме «кирпич и раствор». Липиды также были обнаружены в следах гекконов, внутри щетинок и на поверхности ткани щетинок. Липиды играют важную роль в различных биологических системах, включая эпидермис млекопитающих, рептилий и птиц, где они, помимо прочего, выполняют функцию водного барьера. Также было доказано, что липиды играют роль в самосборке белков в биссусных нитях мидий и, таким образом, способствуют адгезивным свойствам мидий.
Липиды потенциально могут сделать белковую ткань щетинки гидрофобной, что часто является основой для самоочищающихся структур, и тем самым могут поддерживать такие свойства щетинок геккона.
Чтобы детальнее исследовать химическую структуру липидного слоя щетинок геккона (вид Gekko gecko), ученые использовали спектроскопию методом NEXAFS (X-ray absorption near edge structure). Для спектроскопии NEXAFS поверхность облучают рентгеновскими лучами при энергиях фотонов вблизи края ионизации. Испускаемые электроны детектируются. Когда энергия падающего фотона находится в резонансе с переходом электронов основного уровня на незанятые молекулярные орбитали вблизи края ионизации, выход электронов увеличивается. Интенсивность также зависит от относительной ориентации входящих рентгеновских лучей и соответствующего дипольного момента перехода, что позволяет исследовать молекулярное выравнивание и ориентацию. Конкретно в этом труде рассматривались молекулярные орбитали вблизи K-края* углерода и K-края азота.
K-край* — в рентгеновской спектроскопии это внезапное увеличение поглощения рентгеновских лучей, происходящее, когда энергия рентгеновских лучей чуть выше энергии связи самой внутренней электронной оболочки атомов, взаимодействующих с фотонами.
Так как регистрируются только электроны с энергией выше определенного порога, которые исходят от молекул вблизи поверхности материала, NEXAFS-спектроскопия является очень поверхностно-чувствительным методом с глубиной зондирования примерно 5–10 нм.
Результаты исследования
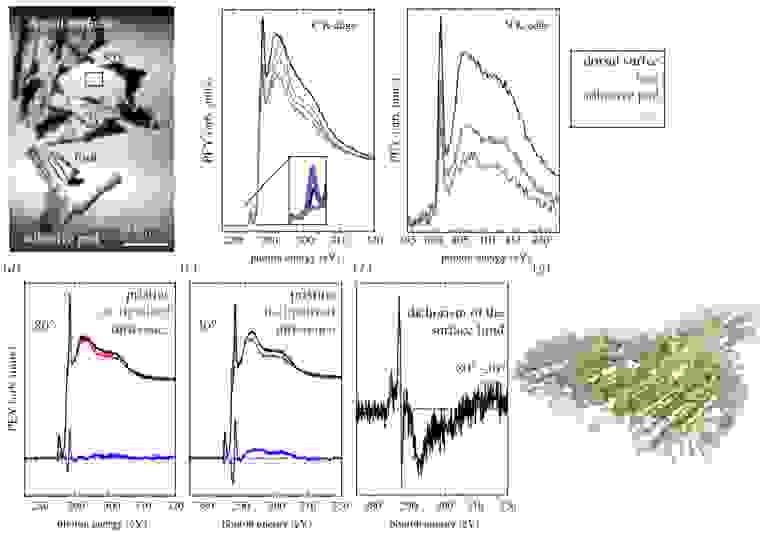
Изображение №2
На изображении №2 показаны спектры NEXAFS углеродного K-края и азотного K-края щетинок геккона и трех типов контрольных тканей (дорсальная чешуя тела, дорсальная чешуя стопы и чешуя века).
Преимущество использования метода визуализации NEXAFS-спектроскопии в контексте анализа тканей состоит в том, что микроскоп основан на магнитном соленоиде, передающем специальный выход электронов, и визуализации канальной пластины для обнаружения электронов, что обеспечивает зондирование изогнутых непроводящих образцов без спектральных искажений.
Спектры NEXAFS на 2b были извлечены из данных изображения путем построения графика зависимости PEY (partial electron yield, т. е. частичный выход электронов) от энергии фотонов для четырех областей интереса (ROI от regions of interest). Все спектры демонстрируют алифатический σ*-резонанс вблизи 292 эВ, а также C=C и C=O π*-резонансы вблизи 285 эВ и 288 эВ соответственно. Эти спектральные особенности ожидаются для образцов кожи геккона, содержащих сложные биомолекулы, а формы спектров напоминают формы для липидов и белков. При сравнении состава липидов и белков было выявлено, что белки имеют большее содержание С=О по сравнению с липидами. Для образцов ткани геккона интенсивность C = O несколько меньше на поверхности щетинок, что указывает на несколько большее количество липидов, покрывающих структуру белка, по сравнению с другими образцами тканей.
π* резонансы C=C относительно сильны в местах, связанных с дорсальной частью стопы, и слабее на веке и дорсальной чешуе тела. Это может свидетельствовать о том, что белки чешуи и щетинок тыльной стороны стопы состоят из большего количества ароматических аминокислот (например, тирозина). На 2c показаны NEXAFS спектры азота до K-края.
Спектры азота содержат амидный π*-резонанс около 401 эВ, обычно обнаруживаемый в спектрах белков, наряду с σ*-резонансами выше 405 эВ, относящимися к N-C и N-H-содержащим группам в основной и боковых цепях белков. Липиды также могут содержать соединения азота, а некоторые липиды, такие как сфингомиелин, также содержат амидные группы, которые могут влиять на спектры амидов белка. Поэтому, чтобы определить конкретный вклад липидов в спектры азота, были собраны эталонные измерения для образцов, которые были лишены липидов. Образцы с удаленными липидами не показали существенной разницы по сравнению с тестируемой тканью геккона, и поэтому можно исключить значительный вклад липидов в сигнал амида.
В результате спектральные особенности, вероятно, связаны с β-белками в ткани геккона. Глубина зондирования при использовании NEXAFS составляет 5 нм. Поскольку обнаружен сигнал азота, NEXAFS, очевидно, проникает через липидный слой в белковую матрицу спатулы. Это означает, что слой липидов на поверхности спатулы должен быть тоньше, чем глубина взятия пробы, и иметь толщину до 5 нм.
Предполагая, что липиды сохраняются как функциональный слой между циклами линьки, возникает вопрос, как липидный слой пополняется во время передвижения. В предыдущих исследованиях сообщалось о наличии липидов в следах гекконов. При шаге 10 Гц для бегущего геккона поверхность спатулы должна быть повторно покрыта в течение 100 мс.
Для покрытия поверхности спатулы площадью примерно 2 × 105 нм2 путем поверхностной диффузии через стержень спатулы диаметром 100 нм и при коэффициенте диффузии примерно 10–8 см2/с для замены липидного слоя потребуется примерно 250 мс, что слишком медленно.
Также возможно, что процессы как поверхностной, так и объемной диффузии играют роль и обеспечивают надежный механизм пополнения липидов для адгезивной системы геккона. В частности, учитывая сильную зависимость процесса диффузии от дополнительных факторов окружающей среды, высокая температура плавления вовлеченных липидов, которая может серьезно препятствовать транспорту липидов через поверхность стержня шпателя.
Как структуру, так и ориентацию липидного покрытия можно определить с помощью NEXAFS спектроскопии с угловым разрешением. Для этого ученые провели сравнение щетинок спатулы гекконов с и без липидного слоя (NEXAFS 80° на 2d, NEXAFS 30° на 2e).
Как видно из отрицательной полярности резонанса около 286.7 эВ для обоих углов, по отношению к общему содержанию углерода в липидном покрытии присутствует меньше групп C-H, чем в нижележащем кератиновом слое, тогда как C=O и C- C выше по интенсивности на природном образце. Относительное количество связывания C-H и C=O хорошо согласуется с гипотезой о том, что липиды удаляются во время движения, поскольку липиды имеют более высокое относительное количество групп C-H и меньшее количество групп C=O по сравнению с белками.
Ориентацию и порядок молекулярных связей можно определить по изменениям поглощения рентгеновских лучей при изменении угла образца по отношению к падающим рентгеновским лучам. Используя разность спектров на 2a и 2b, можно определить угловую зависимость (т. е. дихроизм NEXAFS) липидного слоя (2f).
Принимая во внимание относительную ориентацию входящих рентгеновских лучей и молекулярных орбиталей можно определить ориентацию липидов. Разностный спектр липидного слоя на 2f демонстрирует положительный дихроизм для резонансов π*(C=C) и σ*(C−H). Разностный спектр демонстрирует высокую степень упорядочения алкильных цепей и карбоксильных фрагментов на поверхности, как и ожидалось для упорядоченного липидного слоя. Положительный дихроизм для связей C=C и C-H с отрицательным дихроизмом для связей C=O и C-C является признаком вертикальной ориентации липидных алкильных цепей. Схематическая модель структуры липидов относительно геометрии белкового остова показана на 2g. Основываясь на вертикальной ориентации липидов, средней длине липидов около 2 нм и максимальной толщине слоя 5 нм, можно сделать вывод, что липидная пленка состоит из трех слоев.
Ученые отмечают, что на данный момент можно строить лишь предположения касательно функций липидного слоя. Однако межфазные липиды могут иметь несколько потенциальных преимуществ для адгезии. К примеру, липидный слой может участвовать в поддержании гидратации белков щетинок и спатулы, окружая белковую ткань и тем самым уменьшая испарение воды. Ранее было показано, что высушенные щетинки геккона прилипают сильнее, когда они находятся в условиях высокой влажности (гидратации). Возможно, это связано с более низким модулем упругости щетинок в гидратированном состоянии, что приводит к усилению образования контактов с наноразмерной шероховатостью. Этот эффект может быть подобен роли липидов в щетинках насекомых, где липиды предотвращают высыхание резилина (резиноподобного белка со свойствами гидрогеля, присутствующего на кончиках щетинок).
Полярность поверхности может играть важную роль в адгезии, а липиды могут придавать белковым поверхностям спатулы гидрофобность. Взаимодействие между гидрофобными поверхностями могут быть достаточно сильны. Учитывая место обитания гекконов (влажные регионы), то поверхности, по которым они перемещаются, являются достаточно влажными (к примеру, листья растений). Кроме того, гидрофобный массив щетинок может приводить к обезвоживанию субстратов, покрытых водой, за счет использования захваченных пузырьков воздуха между щетинками. Самоочищающееся свойство щетинок, особенно в случае гидрофильных частиц пыли, и их способность быстро удалять воду с влажных подушечек пальцев лап могут поддерживаться именно гидрофобными свойствами массивов щетинок.
Основной вывод данного исследования заключается в том, что щетинки геккона покрыты нанометровым слоем упорядоченных липидов. Липиды ориентированы вертикально, что указывает на высокую плотность и выравнивание. Но для более полного описания функций этого слоя требуются дополнительные исследования.
Для более детального ознакомления с нюансами исследования рекомендую заглянуть в доклад ученых и дополнительные материалы к нему.
Эпилог
В рассмотренном нами сегодня труде ученые установили, что поверхность лапок гекконов покрыта тонким слоем липидов. Если раньше считалось, что адгезионные свойства гекконов связаны исключительно с наличием на их подушечках пальцев специальных щетинок, то сейчас было высказано предположение о том, что и липиды играют не последнюю роль.
Как говорят сами ученые, полноценное понимание физических и химических процессов, которые задействованы в умении гекконов прилипать буквально ко всему, являются крайне важной информацией для будущих разработок адгезионных материалов. Конечно, им предстоит проделать еще немало работы, но уже сейчас они видят огромный потенциал своего открытия.
Немного рекламы
Спасибо, что остаётесь с нами. Вам нравятся наши статьи? Хотите видеть больше интересных материалов? Поддержите нас, оформив заказ или порекомендовав знакомым, облачные VPS для разработчиков от $4.99, уникальный аналог entry-level серверов, который был придуман нами для Вас: Вся правда о VPS (KVM) E5–2697 v3 (6 Cores) 10GB DDR4 480GB SSD 1Gbps от $19 или как правильно делить сервер? (доступны варианты с RAID1 и RAID10, до 24 ядер и до 40GB DDR4).
Dell R730xd в 2 раза дешевле в дата-центре Maincubes Tier IV в Амстердаме? Только у нас 2 х Intel TetraDeca-Core Xeon 2x E5–2697v3 2.6GHz 14C 64GB DDR4 4×960GB SSD 1Gbps 100 ТВ от $199 в Нидерландах! Dell R420 — 2x E5–2430 2.2Ghz 6C 128GB DDR3 2×960GB SSD 1Gbps 100TB — от $99! Читайте о том Как построить инфраструктуру корп. класса c применением серверов Dell R730xd Е5–2650 v4 стоимостью 9000 евро за копейки?
