Когда морщины на пользу: кинематика хобота слона
Если взглянуть на некоторые творения эволюции беглым взглядом, то может показаться, что у нее отличное чувство юмора. Однако любая «странность» странной не является, так как выполняет определенную функцию, которая в той или иной степени влияет на выживаемость вида. В то же время, смесь любопытства и прагматичности заставляет нас с большим интересом изучать любые необычные особенности представителей флоры и фауны. Ученые из Технологического института Джорджии (США) не исключение. Они предположили, что подвижность хобота слона зависит не только от мышечной ткани, но и от своеобразной морфологии кожи, а потому решили проверить эту гипотезу. Какие наблюдения были проведены, что они показали, и как полученные знания могут быть применены на практике? Ответы на эти вопросы мы найдем в докладе ученых. Поехали.
Основа исследования
Думая о слонах, мы вспоминаем о том, что это крупные млекопитающие с большими ушами и длинным хоботом. Это, конечно, крайне скудное описание слона, но упрекать человека в том, что он замечает столь ярко выраженные особенности слоновьей внешности, не стоит. Несмотря на некую комичность, эти особенности имеют вполне логичное объяснение и практическое применение. Уши, к примеру, позволяют слонам регулировать температуру тела, так как оснащены огромным количеством кровеносных сосудов.Водные, грязевые и даже пыльные ванны — спасательное средство от вездесущий жары.
Хобот, как и многое другое у слонов, не появился у них изначально, а стал результатом многолетней эволюции, объединившей нос и верхнюю губу слоновьих предков в функциональный «нюхательно-хватательный» манипулятор длиной 1.5 метра. Слоны не пьют воду через хобот, как это часто изображается в мультфильмах, но используют его как черпак, набирая воду, а потом выливая в рот. К слову, слон может за один раз выпить порядка 90 литров воды. Несмотря на свои габариты, слоны отлично плавают, практически полностью погружаясь под воду, выставив на поверхность лишь хобот.
Короткое, но познавательное видео о хоботе слона.
С помощью хобота слоны могут не только обонять и дышать, но и тактильно взаимодействовать с окружающим миром, поднимая предметы с земли, срывая плоды с деревьев и т. д. Внешне хобот слона не одинаков со всех сторон, а имеет два типа кожи. С тыльной стороны хобота кожа достаточно нежная и чувствительная, что помогает при хватании предметов. С внешней стороны кожа толстая и грубая, позволяя слонам буквально таранить хищников или пробираться сквозь заросли.
В отличие от скелетно-мышечной системы с костями и суставами хобот представляет собой мышечный гидростат: бескостную структуру, состоящую из плотно упакованных мышечных волокон, обернутых кожей. Подобные системы не уникальны для слонов и встречаются в природе достаточно часто (язык млекопитающих, щупальца осьминога и т. д.). Различная ориентация мышечных волокон позволяет выполнять различные движения относительно оси хобота, включая крен, тангаж и рыскание.
Любопытный факт: самыми крупными мускульными гидростатами считаются язык синего кита (массой в 2600 кг) и щупальце гигантского кальмара (длиной 12 м).
Вполне логично, что именно мышцы в таких системах привлекают больше всего внимания ученых, особенно в рамках бионики и робототехники. Однако и кожа играет важную роль. В конце XVIII века слонов первоначально классифицировали как толстокожих, то есть животных с толстой кожей. Действительно, африканский слон весом 3500 кг имеет измеренную площадь поверхности кожи 20.7 м2, что примерно в 10 раз превышает площадь поверхности кожи человека (2.04 м2). Как и кожа всех млекопитающих, кожа слона действует как физический и иммунный барьер, блокируя потенциальные опасности окружающей среды, такие как вирусы, бактерии, ультрафиолетовый свет и токсины. Кожа слона также защищает тело от мелких порезов и ссадин и лишена потовых желез, чтобы уменьшить потерю воды. Когда слоны покрываются грязью, морщины на их коже удерживают влагу, что позволяет им лучше охладиться. Стоит отметить, что африканские слоны более морщинистые и обладают большими ушами, чем их азиатские родственники, именно из-за разных климатических условий обитания. Авторы рассматриваемого нами сегодня труда считают, что кожа слона играет очень важную роль не только в терморегуляции, но и в подвижности его конечностей.
Результаты исследования
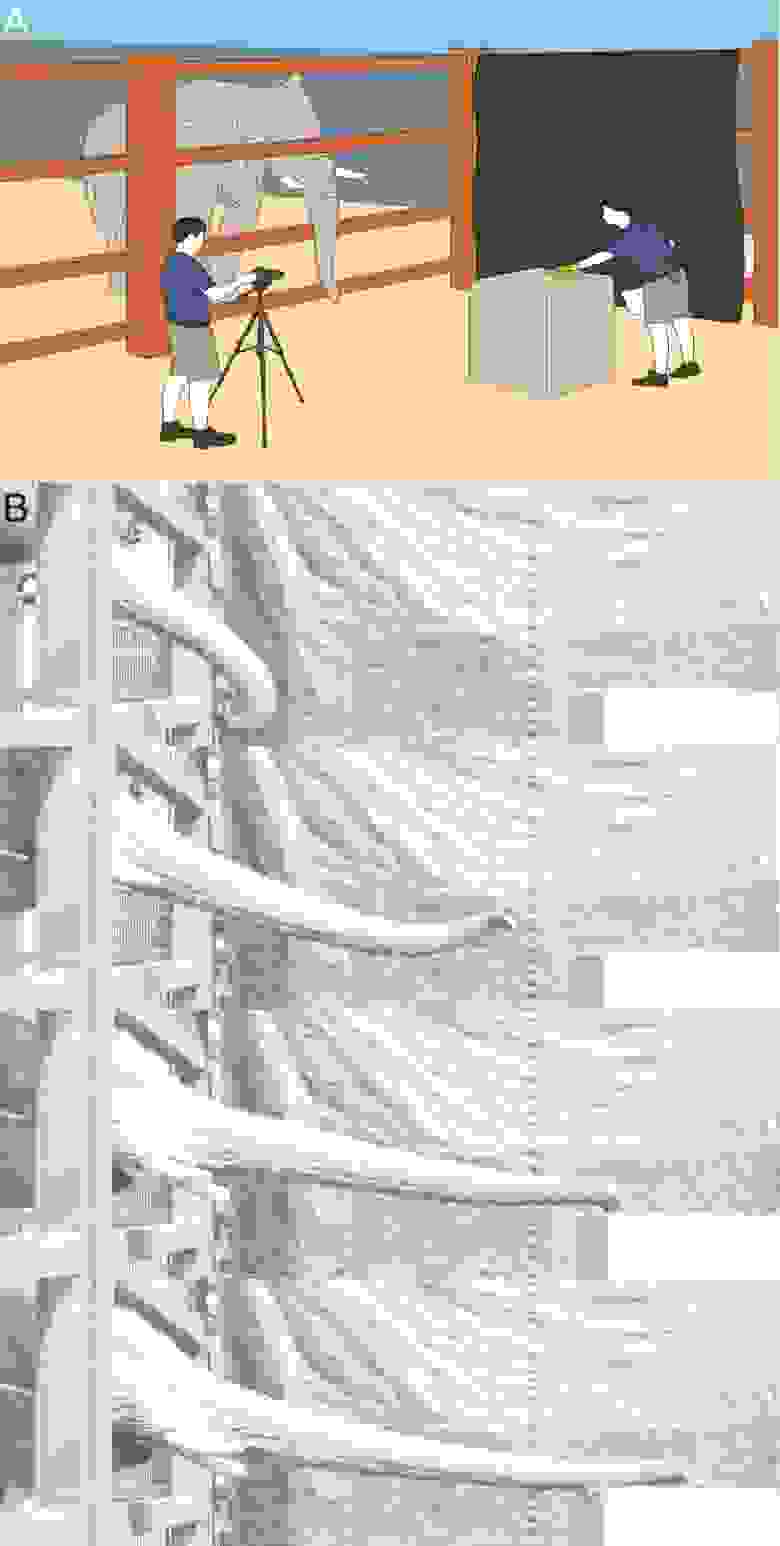
Для данного исследования ученые провели ряд наблюдений за самкой и самцом африканского слона, которые пытались хоботом дотянуться до объекта, расположенного на определенном расстоянии (1A). В ходе этих наблюдений фиксировались изменения кожи хобота.
Изображение №1
Движение, которые слон совершал в попытке дотянуться до вкусняшки, можно разделить на три фазы: вис (свисание), подъем и удлинение (1B). Когда слон отдыхал, то длина хобота в висячем положении составляла 126 см (самка) и 141 см (самец). Исходя из этой исходной длины, было проведено описание деформации хобота с точки зрения напряжения, отношения изменения длины во время удлинения к его первоначальной длине. Например, деформация 0.2 указывает на то, что хобот увеличился в длину на 20% от длины его висячего положения, а деформация -0.2 указывает на то, что он сократился на 20%.
Изображение №2
Первое растяжение произошло, когда хобот поднялся в горизонтальное положение. Во время этого подъема напряжение в хоботе увеличилось на 0.20 ± 0.045 (n = 7) у самки и на 0.12 ± 0.012 (n = 3) у самца. Таким образом, поднятие хобота подготавливает его к вытягиванию, вызывая мышечные сокращения, которые существенно его удлиняют. Хобот удлинялся дальше во время тянущегося движения, создавая конечное напряжение 0.25 для самки и 0.13 для самца (изображение №2). Каждое испытание состояло как из фазы подъема, так и из фазы удлинения, комбинация которых имела продолжительность 2 ± 0.15 с (n = 10).
Видео №1: самка слона пытается дотянуться до объекта на расстоянии 200 см.Видео №2: самец слона пытается дотянуться до объекта на расстоянии 200 см.
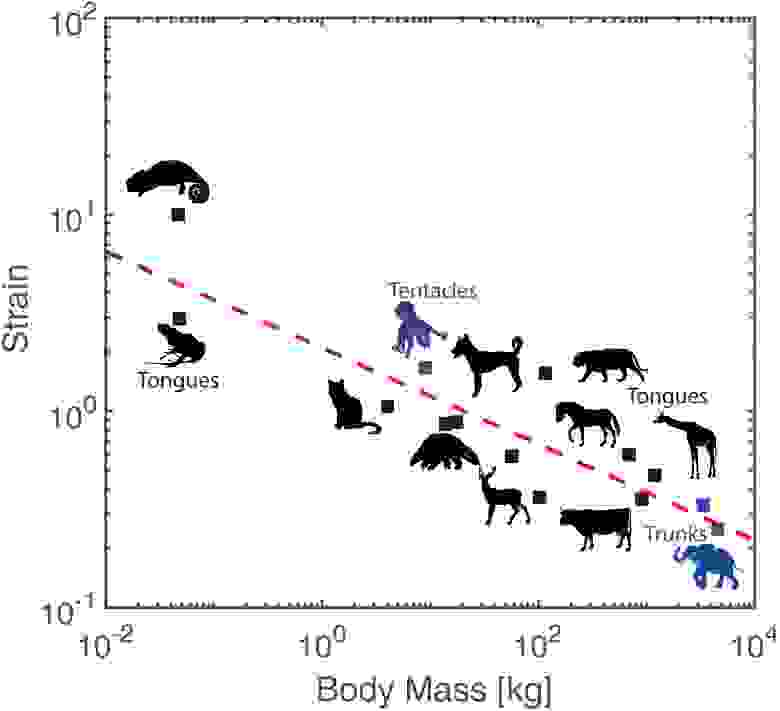
Необходимо было также установить, чем напряжение хобота слона отличается от других мышечных гидростатов. Для этого были собраны данные по гидростатам 13 типов (язык хамелеона, язык разных млекопитающих, щупальца головоногих и т. д.). На графике выше показана зависимость между массой тела (M) и максимальной осевой деформацией (εA). Пунктирная линия показывает наилучшее приближение по степенному закону:

где М выражено в килограммах, а R2 = 0.68 — логарифмический коэффициент детерминации. Степенной закон дает интерполированные значения, которые обычно хорошо согласуются с экспериментальными измерениями. Например, осьминоги (9 кг) демонстрируют напряжение 2.66, что близко к интерполяции 2.57. Самки (3400 кг) и самцы (4550 кг) слонов имеют деформации 0.25 и 0.13 соответственно с интерполяцией 0.15 и 0.07. Очевидно, что напряжение уменьшается с увеличением размера тела, и эта тенденция сохраняется даже у слонов разного пола. Единственным «уникумом» является хамелеон (0.047 кг), который демонстрирует деформацию 10.0, что намного больше, чем интерполяция 5.37. Связано это с тем, что хамелеон развил в своем языке серию из 10 концентрических оболочек, которые позволяют ему вытягиваться как телескоп.
Изображение №3
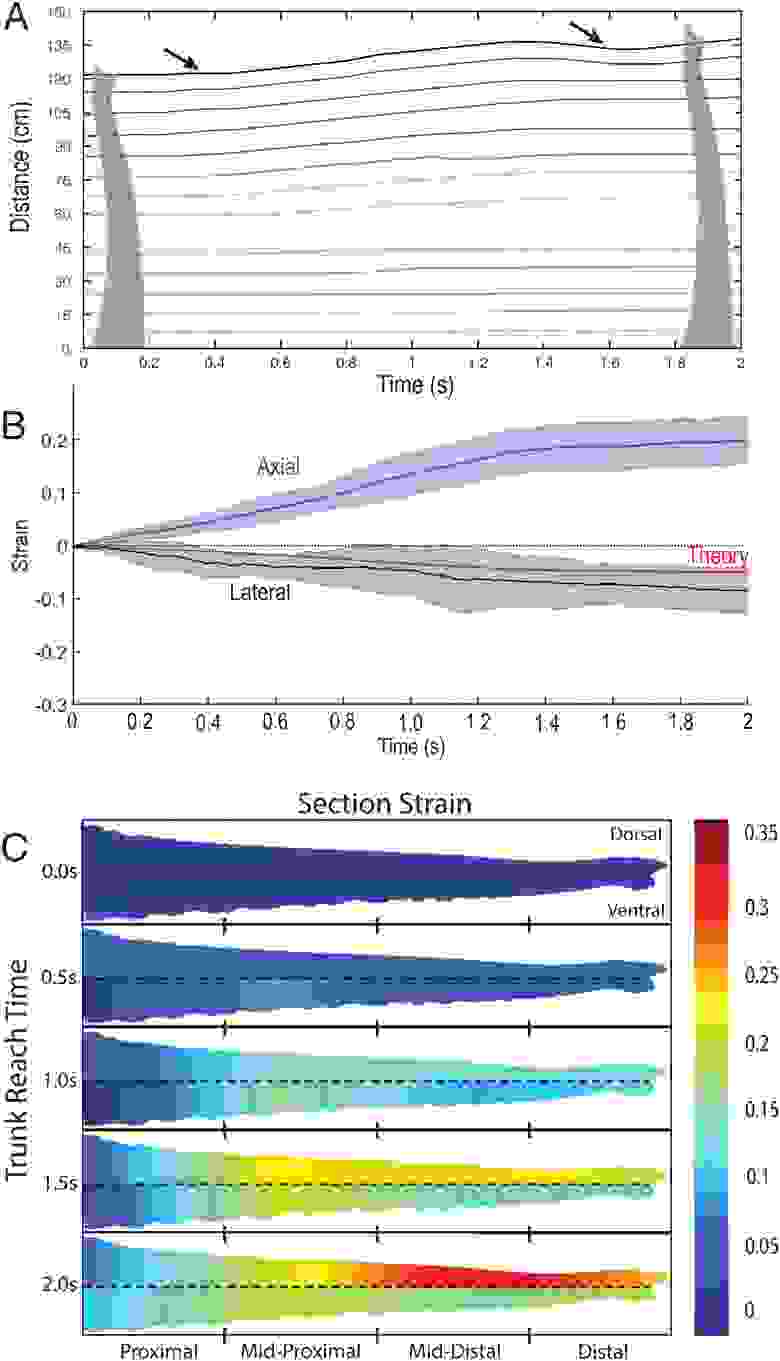
У осьминогов и большинства языков млекопитающих удлинение происходило равномерно по всей длине придатка. Однако в хоботах слонов удлинение происходило волнами растяжения, которые начинались с кончика и двигались к основанию. На 3А показан ход во времени положения 14 равноотстоящих точек вдоль хобота слонихи.
Как показано черными стрелками, были сгенерированы две волны, одна с началом при t = 0.4 с, а другая с началом при t = 1.6 с (3А). Предположительно, первая волна произвела недостаточное удлинение, чтобы достичь объекта, поэтому слон «запустил» вторую волну. В отличие от первой волны, вторая волна дошла только до середины хобота, которая, по-видимому, отмечает предел удлинения хобота.
Как самец, так и самка показали по крайней мере один пик скорости деформации, соответствующий первой из двух бегущих волн, описанных выше. Самец слона имел максимальную скорость деформации 0.25 с–1, приходящуюся в момент времени t = 0.2 с. Точно так же у самки слона максимальная скорость деформации составляла 0.2 с–1 в момент времени t = 0.85 с. Похожая картина наблюдается и у щупалец кальмара, когда скорость деформации также высока.
Одним из ограничений гидростатических органов является то, что они следуют закону сохранения массы. Поскольку объем органа фиксирован, продольное растяжение должно сопровождаться сокращением в латеральном направлении. Подобная картина описывает как щупальца кальмаров, так и язык млекопитающих, но хобот слона, содержащий два полых носовых хода, еще не оценивался в рамках этой модели.
Следовательно, модель гидростатической системы хобота включает данные по внутреннему и внешнему размерам хобота, полученные с помощью КТ. Основываясь на измеренной осевой деформации, можно было предсказать боковую деформацию для слонихи, а потом провести сравнение с экспериментальными результатами (3B).
Видео №3: отслеживание положения точек на тыльной и внешней сторонах хобота. Зеленые точки — изначальное положение, красные — положение, когда слон тянется к объекту.
Основываясь на средней осевой деформации вдоль хобота 0.199 ± 0.045 (n = 7), была рассчитана латеральная деформация -0.039, что на 0.046 меньше по величине, чем наблюдаемое значение в -0.086 ± 0.043 (n = 7). Более высокие значения в моделировании могут быть связаны с ошибкой аппроксимации: в модели поперечное сечение хобота представляет собой круг по всей его длине, а в реальности хобот округлый с внешней стороны и слегка сплющенный с тыльной.
Данная модель показала, что наблюдаемая боковая деформация согласуется с уменьшением объема на 0.18 л, что составляет 0.2% от объема хобота. Из этого следует, что хобот слона действительно следует закону сохранения массы.
Также было обнаружено, что хобот слона демонстрирует поразительные различия в вентральной и дорсальной деформациях. Чтобы лучше описать расположение наибольшего вентрально-дорсального несоответствия, хобот был разделен на четыре секции длиной 30 см: проксимальный, среднепроксимальный, среднедистальный и дистальный отделы.
Видео №4: Осевые деформации дорсального и вентрального отдела хобота показаны в виде последовательности тепловых карт, отображающих изменение длины с течением времени.
Тепловая карта на 2C показывает, что через 2 секунды удлинения дорсальная среднедистальная часть хобота (30–60 см от кончика) испытывает напряжение до 0.35, в то время как соответствующий вентральный отдел — 0.18. Аномальная разница составляет 0.17, что эквивалентно дополнительному смещению 5.1 см в дорсальном отделе. Это смещение существенно по сравнению с другими областями хобота, где разница между дорсальной и вентральной областями на порядок меньше. Например, проксимальный отдел (от 90 до 120 см) имеет дорсальную и вентральную деформации 1.10 и 1.12 соответственно, т. е. разница составляет всего лишь 0.5 см.
Ввиду всех вышеописанных данных возникает вопрос — какая польза в том, что слон тянется хоботом именно дорсальной частью хобота? Сначала ученые предположили, что разница в деформации была связана с тем, что дорсальный отдел хобота движется по кривой и, следовательно, более длинной траектории для достижения цели. Однако 18 из 20 горизонтально расположенных точек на дорсальной и вентральной областях показали незначительные значения кривизны, о чем свидетельствуют их значения P > 0.05. Другие две точки хоть и имели P менее 0.05, но показатели кривизны были столь малы, что их вполне можно игнорировать. Более того, эти области не соответствовали срединно-дорсальному отделу, где наблюдалась асимметрия.
Вместо этого ученые предположили, что дорсальный конец хобота растягивается дальше, что помогает кончику хобота наклоняться вперед, чтобы достичь объекта, как показано в момент времени t = 0.95 с на 1B. Наблюдаемая асимметрия не является случайным наблюдением, а является повторяющейся чертой всех испытаний как для самцов, так и для самок слонов. Чтобы найти объяснение данной асимметрии, ученые решили провести наблюдения за анатомией и мышц, и кожи хобота (взятого из музея).
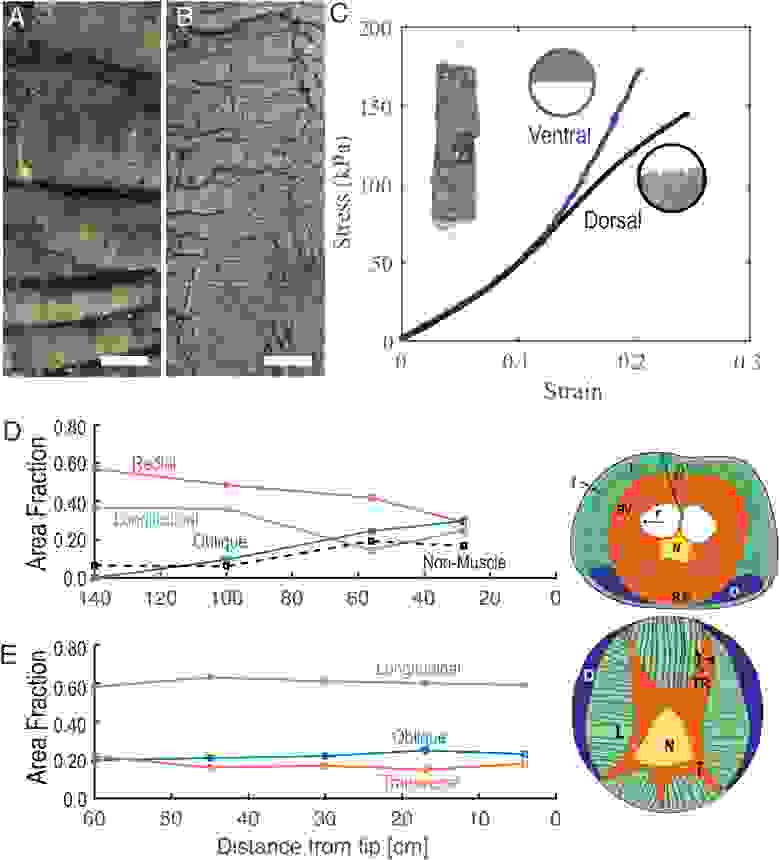
Изображение №4
Аномальное растяжение дорсального туловища согласуется с растяжимостью дорсальной кожи. Замороженный хобот слона имеет дорсальную кожу, состоящую из ряда перекрывающихся складок длиной 4 см. Напротив, вентральная кожа состоит из морщин меньшего размера (от 1 до 2 см).
На 4C показаны измерения напряжения-деформации дорсальной и вентральной кожи, взятые из проксимального отдела основания хобота. Аналогичная картина наблюдалась и во время экспериментальных наблюдений.
В частности, дорсальный и вентральный отделы имеют сопоставимую эластичность в линейном режиме с модулем Юнга 470 ± 15 кПа (n = 3) для дорсального и 483 ± 13 кПа (n = 3) для вентрального отдела (4C). Линейный режим реализуется в пределах деформаций от 0 до 0.15. Этот режим деформации близко соответствует наблюдаемым напряжениям в экспериментах по удлинению, когда спинная и брюшная секции выравниваются (до момента времени 1.0 с на 2B).
Выше этой деформации разная геометрия морщин и складок влияет на их эластичность. Нелинейный режим испытаний на растяжение показал, что дорсальная область поддерживает модуль Юнга, сравнимый с ее линейным режимом, 670 ± 65 МПа (n = 3), в то время как вентральная область демонстрирует более жесткий модуль Юнга 1.19 ± 0.12 МПа (n = 3).
Отношение модулей Юнга составляет 1.77, что близко соответствует 1.75 — соотношению дорсальной и вентральной деформаций в нелинейном режиме экспериментов по удлинению. Из вышеописанных данных следует, что удлинение хобота слона осуществляется не только за счет мышц, но и за счет свойств их кожи.
Видео №5: Видео анализа методом цифровой корреляции изображений (DIC), выполненного на растянутой коже слона.
Подтвердив участие кожи в удлинении хобота, ученым осталось выяснить, насколько сильно кожа влияет на этот процесс.
Ранее было измерено, что жесткость мышц хобота слона составляет 938 кПа, что делает кожу слона не более чем в 1.27 раза более жесткой, чем мышечная ткань. Если хобот растягивается, необходимая энергия деформации мышц и кожи будет пропорциональна EV, где E — модуль Юнга, а V — вытесненный объем. Используя собранные данные относительно деформации хобота, ученые рассчитали отношение λ энергии растяжения кожи к энергии растяжения мышц.
На расстоянии 20 см от кончика кожа имеет в λ = 13.2 раза большую энергию деформации, чем соответствующая мышца. Такая внушительная разница возникает из-за того, что ноздри преобладают во внутреннем объеме на кончике хобота, что увеличивает отношение объема кожи к объему мышц. И наоборот, на расстоянии 110 см от кончика хобота кожа имеет только 0.36 от энергии напряжения мышцы, потому что ноздри занимают гораздо меньшую часть поперечного сечения. Эти расчеты объясняют, почему наблюдается асимметрия только в одном отделе хобота (около кончика, а не у основания).
Асимметрия растяжения хобота соответствует его внутренней анатомии. На 4D показаны пропорции трех типов мышц в зависимости от расстояния от кончика хобота: продольные мышцы (зеленый), радиальные или поперечные мышцы (оранжевый) и косые мышцы (синий). Для полноты картины ученые также учли и немышечные компоненты хобота (носовая полость, нервы, кожа и кровеносные сосуды), отмеченные пунктирной линией на 4D.
Косые мышцы, используемые для рыскания хобота влево/вправо, обнаруживаются только в вентральной области с возрастающей долей от 0% у основания и до 25% на конце (4D). Хотя рысканье отсутствует при одноосном растяжении, наличие косых мышц в вентральном отделе может ограничивать растяжение, особенно в средне-дистальных отделах, где косые мышцы составляют наибольшую часть поперечного сечения.
Асимметричное распределение косых мышц кажется уникальным для хобота слона. Как показано на 4Е, другие гидростатические органы, такие как щупальца осьминога вида Octopus bimaculoides, обычно радиально-симметричны. Более того, пропорция продольных, поперечных и косых мышц постоянна от основания до кончика, ~60%, 20% и 20% соответственно.
Для более детального ознакомления с нюансами расследования рекомендую заглянуть в доклад ученых и дополнительные материалы к нему.
Эпилог
Из вышеописанных результатов моделирования и экспериментальных наблюдений следует, что слоны способны вытягивать хобот на 10–20%, чтобы дотянуться до удаленных объектов. Этот процесс происходит телескопически с одной или двумя волнами растяжения, направленными от дистального конца к проксимальному основанию хобота. Было обнаружено, что дорсальный отдел хобота более гибкий, нежели вентральный. И вентральная, и дорсальная части равномерно удлиняются в первой половине процесса дотягивания. Во второй половине процесса дорсальный среднедистальный отдел продолжает растягиваться на 15%, в то время как соответствующий вентральный отдел остается фиксированной длины.
Такая асимметрия в растяжении близка к поведению кожи, которое ученые наблюдали во время тестов с замороженным хоботом слона. Было обнаружено, что при высоких нагрузках жесткость кожи тыльной стороны хобота в 1.77 раза больше, чем наружной стороны. Из-за этого подвижность хобота на дистальном конце становится меньше, ввиду наличия ноздрей.
Суть заключается в том, что на внешней стороне хобота, где много складок, кожа намного более эластичная (на 15%), чем слегка морщинистая тыльная сторона. Хобот слона отличается от других бескостных, наполненных мышцами придатков, встречающихся в природе (щупальца кальмара, язык млекопитающих и т. д.). Вместо того чтобы вытягиваться равномерно, слон телескопически вытягивает свой хобот, как зонтик, постепенно удлиняя его волнами.
По словам ученых, данное исследование не только позволяет лучше понять то, как устроена анатомия различных обитателей нашей планеты, но и вполне может послужить вдохновением в мягкой робототехнике. Большинство мягких роботов тем или иным образом основаны на движении мышц, а механику кожи часто не учитывают. Однако, как показали результаты этого труда, такое пренебрежение кожей является большой ошибкой. Если в бионике поставлена цель воссоздать какую-то биологическую систему в механическом или электронном виде, то нужно детально изучить все аспекты оригинала, прежде чем приступать к созданию копии.
Немного рекламы
Спасибо, что остаётесь с нами. Вам нравятся наши статьи? Хотите видеть больше интересных материалов? Поддержите нас, оформив заказ или порекомендовав знакомым, облачные VPS для разработчиков от $4.99, уникальный аналог entry-level серверов, который был придуман нами для Вас: Вся правда о VPS (KVM) E5–2697 v3 (6 Cores) 10GB DDR4 480GB SSD 1Gbps от $19 или как правильно делить сервер? (доступны варианты с RAID1 и RAID10, до 24 ядер и до 40GB DDR4).
Dell R730xd в 2 раза дешевле в дата-центре Maincubes Tier IV в Амстердаме? Только у нас 2 х Intel TetraDeca-Core Xeon 2x E5–2697v3 2.6GHz 14C 64GB DDR4 4×960GB SSD 1Gbps 100 ТВ от $199 в Нидерландах! Dell R420 — 2x E5–2430 2.2Ghz 6C 128GB DDR3 2×960GB SSD 1Gbps 100TB — от $99! Читайте о том Как построить инфраструктуру корп. класса c применением серверов Dell R730xd Е5–2650 v4 стоимостью 9000 евро за копейки?
