Запрограммировано ли старение? Разбираем доказательства
Гипотеза о том, что старение запрограммировано весьма популярна среди обывателей, но имеет довольно мало сторонников среди ученых-геронтологов. При этом данная гипотеза кажется весьма логичной, на первый взгляд. Откуда такое расхождение? Может быть, ученые просто находятся в плену своих предубеждений и когнитивных ошибок, не давая зеленый свет смелой гипотезе?
«Ничто в биологии не имеет смысла, кроме как в свете эволюции»
Добржанский Феодосий Григорьевич
Для того, чтобы всерьез воспринимать гипотезу о наличии у абсолютного большинства видов некоей генетической программы старения нужны веские основания. Каким образом эволюция привела к тому, что гипотетическая генетическая программа закрепилась (далее просто Программа), какие преимущества она дает? Какую пользу старение может приносить отдельной особи никто так и не придумал, поэтому вся аргументация сторонников Программы сводится к групповому отбору. Групповой отбор — штука спорная, но давайте сделаем вид, что он существует и обратимся к работе Джоша Миттельдорфа, который является одним из сторонников Программы с довольно серьезным научным бэкграундом. Эта работа хороша тем, что она не заявляет голословно, что программа старения полезна для группы организмов, Миттельдорф приводит результаты симуляции, используя хорошо известную и проверенную модель популяционной динамики Лотки-Вольтерры. Давайте посмотрим на динамику численности популяций хищников и их добычи («не-хищников») в разных сценариях (без старения и когда стареют жертвы):
 Моделирование численности популяций нестареющих хищников и жертв.
Моделирование численности популяций нестареющих хищников и жертв.Когда старения нет минимальная и максимальная численность обоих видов разнится на два порядка и довольно долгое время численность популяции находится возле нуля: малейшая флуктуация — и оба вида исчезнут! А теперь взглянем на результаты симуляции, где присутствует старение «не-хищников» и старые особи представляют более легкую добычу:
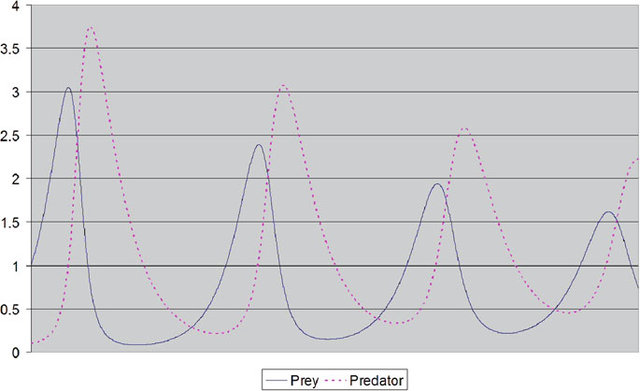 Моделирование численности популяций хищников и стареющих жертв.
Моделирование численности популяций хищников и стареющих жертв.Мы видим, что в этой симуляции динамика существенно стабилизировалась и есть даже некий намек на сходимость — колебания уменьшаются со временем. Это ли не лучшее доказательство наличия Программы?! Прежде чем ответить на этот вопрос, давайте изучим матчасть. А именно модель Лотки-Вольтерры, которую (с небольшими модификациями) использовал Джош Миттельдорф.
Модель Лотки-Вольтерры
«В сущности, все модели неправильны, но некоторые полезны»
Джордж Бокс
В 1925 году Лотка (и независимо от него в 1926 году Вольтерра) предложил модель взаимодействия двух видов типа «хищник — жертва», которая с успехом применяется и сейчас во многих задачах. Модель представляют собой простую систему дифференциальных уравнений и во многом похожа на ставшую печально известной в последнее время эпидемиологическую модель SIR (про которую можно почитать, например, тут).


x — численность жертв (травоядных);
y — численность хищников;
α — скорость роста популяции жертв;
β — вероятность того, что жертва будет съедена хищником при контакте;
γ — 1/γ имеет смысл среднего времени до голодной смерти хищника;
δ — вероятность того, что хищнику хватит еды на дальнейшее размножение.
Оставим пока хищников в покое и рассмотрим уравнение для жертв. Как будет выглядеть динамика их численности при условии, что все хищники куда-то внезапно исчезли (y=0)?
А вот как:
Совершенно непохоже на экспоненциальный рост, не так ли? Современная наука давно объяснила почему так происходит и разработала простую математическую модель, которая точноописывает кривые роста бактерий, дрожжей, инфузорий и даже млекопитающих. И эта модель — рост по логистической кривой (эта кривая изображена черным на рисунке слева). Логистическая модель основана на факте наличия у среды определенной емкости — того максимального количества особей данного вида, которое среда может вместить и прокормить. Наличие такого предела приводит к тому, что скорость роста популяции замедляется по мере приближения к пределу, а при его достижении рост вообще прекращается.
Чем же обусловлено такое замедление? Существуют, как минимум, две группы факторов: механистические и адаптивные (генетические).
К механистическим можно, например, отнести недостаток пространства для размножения. Возьмем, к примеру, паразитическую осу, обитающую в Центральной Америке, которая откладывает яйца (см. рисунок снизу) только на спинках гусениц определенного вида. Очевидно, что максимальный прирост популяции равен количеству гусениц помноженному на количество яиц, которое можно разместить на гусенице. Таким образом ограничивается бесконечный экспоненциальный рост в данном случае. Такая же логика применима и к другим животным: например, существует предел того, сколько гнезд чайки могут разместить в ареале своего обитания, или предел подходящих мест для нереста лососей. Другим чисто механистическим препятствием является голод: если не хватает пищи, то даже если места полно, потомство либо вовсе не родится, либо умрет, не успев продолжить род.

Ко второй группе факторов можно отнести генетические адаптации: у мышей, например, в гипоталамусе обнаружили специальные нейроны, которые «отключают» репродуктивную функцию. Скорее всего, такие нейроны есть и у людей, потому что при экстремальной потере жировой массы женщины временно теряют фертильность. Снижение фертильности при голодании наблюдается и у более простых организмов [еще].
Логистическая функция является решением следующего дифференциального уравнения:

Давайте поймем в чем физический смысл этого уравнения. Пусть x (t) — это функция роста численности некоей популяции от времени, тогда dx/dt — это скорость роста популяции. Понятно, что скорость роста зависит от того, сколько уже особей существует (просто потому что количество детей зависит от количества родителей). Но еще она зависит от множителя (T — x), где T — это некий предел численности популяции, который может обеспечить среда. Когда x = T, то dx/dt = 0, то скорость роста популяции нулевая независимо от того сколько уже особей есть на данный момент). Когда T намного больше x, то функция похожа на экспоненту, но по мере приближения x к T функция становится все более пологой. Это, кстати, похоже на динамику эпидемий и, действительно, рост числа новых случаев COVID-19 неплохо описывается таким простым дифференциальным уравнением. А все почему? Да потому что даже без ограничительных мер вследствие иммунитета (или смерти) у вируса остается все меньше свободных переносчиков для распространения и поэтому скорость распространения падает.
Давайте попробуем решить это уравнение:


Полученная нами функция является той самой логистической функцией, которая применяется для моделирования численности популяций, эпидемий, а также в машинном обучении: в логистической регрессии и искусственных нейросетях (хотя в последних ее уже практически не используют из-за проблемы затухания градиентов, но это совсем другая история). Вот такая вот интересная и полезная функция, которая берет начало из довольно простых соображений.
Итак, чтобы учитывать логистический рост популяции «не-хищников», я переписал первое уравнение Лотки-Вольтерры в следующем виде:

И вот какую динамику популяций мне удалось получить:
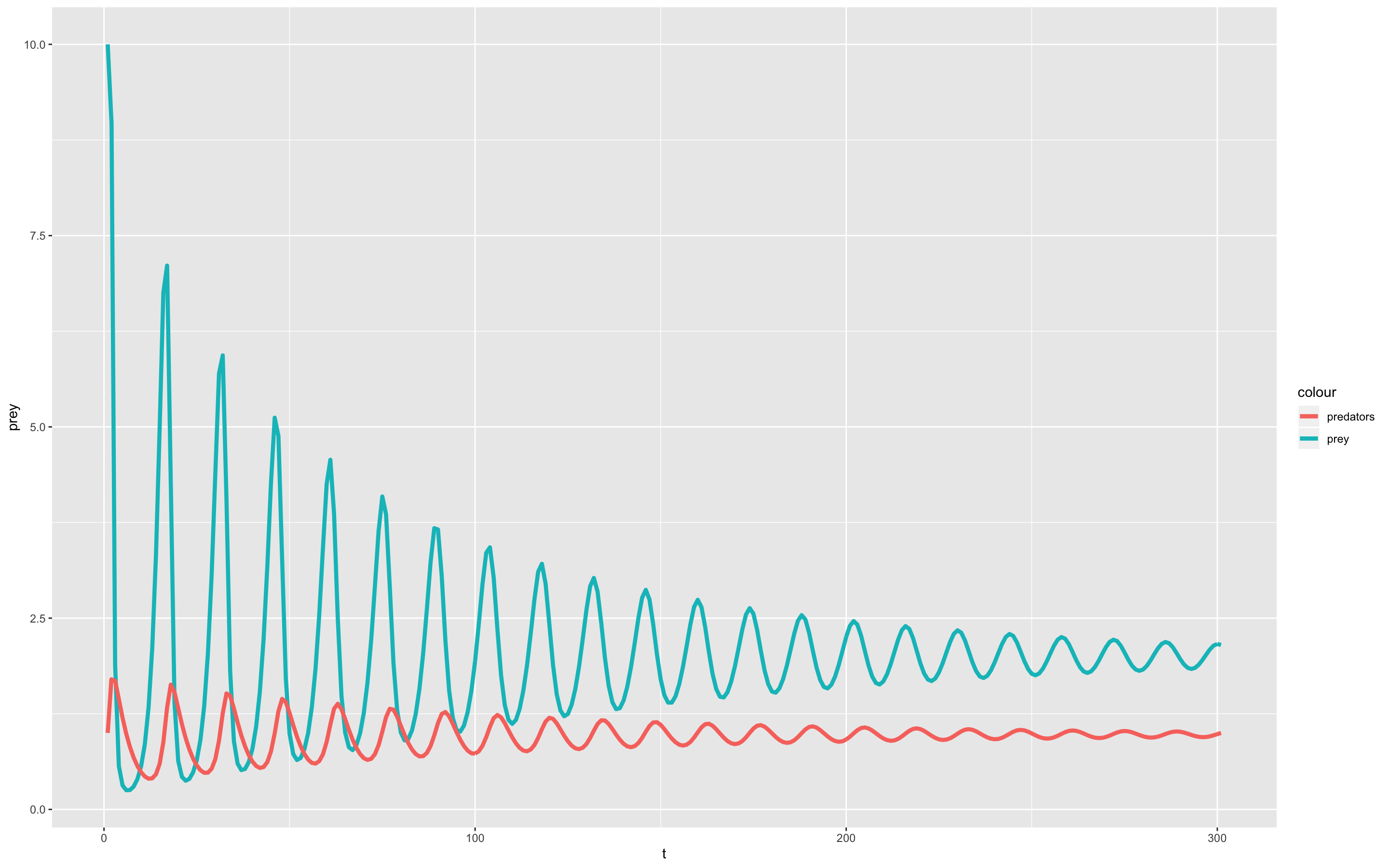
Из графика видно, что замедления репродукции достаточно для стабилизации динамики численности популяций обоих видов! Более того, тут приведена модель другая модифицированная модель, которая учитывает миграцию животных, в рамках которой тоже легко достигается эквилибриум. В общем, нет никакой нужды в программе старения для сглаживания колебаний численности популяций — при использовании более приближенных к реальности моделей колебания исчезают сами по себе.
Есть и другая точка зрения на то, почему эволюция могла привести к закреплению программы старения:
Но в итоге эти виды, когда это преимущество уже закреплялось в генах достаточно большого количества особей, вымирали из-за голода, вызванного перенаселением, от которого такой вид страдал хотя бы один раз в течение миллионов лет эволюции. А избежали вымирания только те виды, в которых групповой отбор закрепил достаточное количество дублирующих механизмов старения, и те, которые научились пережидать «голодные времена» в виде спор или «затаившихся яиц» как нестареющая гидра. https://habr.com/ru/post/405101/
Но спасает ли старение от перенаселения? Давайте рассмотрим простейший пример: пусть у нас есть бактерия, которая умирает от старения через 2 деления. Похожим образом стареют пекарские дрожжи Saccharomyces cerevisiae: от материнской клетки отпочковывается дочерняя клетка, которой достаются самые хорошие органеллы, а все плохие органеллы и клеточный мусор остаются в материнской клетке, которая через несколько десятков таких делений умирает. У дрожжей есть еще хронологическое старение, но это отдельная история.
Так как же будет выглядеть рост популяции в нашей простейшей модели очень быстрого старения? А вот как (см. рисунок внизу для пояснения):
1 бактерия
2 бактерии
4 бактерии — 1 умершая от старости = 3
6 бактерий — 1 умершая от старости = 5
10 бактерий — 2 умерших от старости = 8
16 бактерий — 3 умерших от старости = 13
…
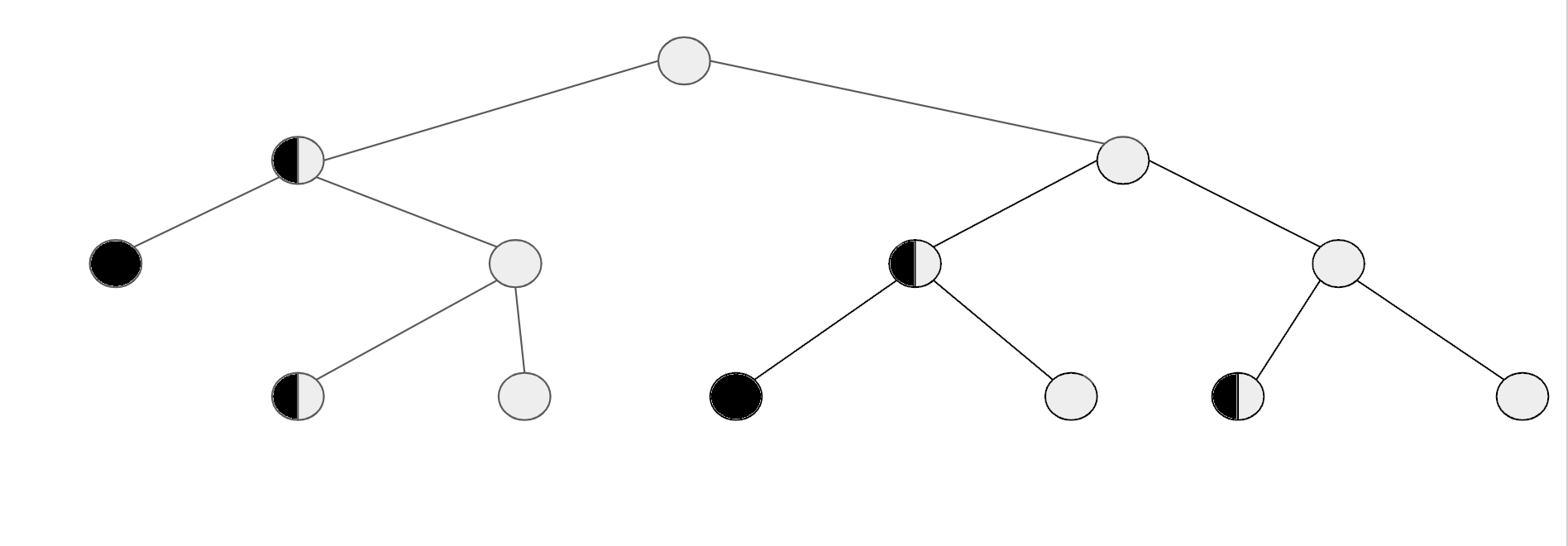 Светлые кружочки — молодые бактерии, наполовину черные — старые, полностью черные — мертвые
Светлые кружочки — молодые бактерии, наполовину черные — старые, полностью черные — мертвые1 2 3 5 8 13… Да это же последовательность чисел Фибоначчи (каждое число равно сумме двух предыдущих)! Но это в свою очередь означает, что старение никак не ограничивает экспоненциальный рост популяции, потому что последовательность Фибоначчи растет по экспоненте. Поэтому эволюция никак не могла прийти к программе старения с этой целью, но она вполне могла прийти к механизмам ограничения размножения, что ученые и обнаружили у мышей.
Это, конечно, не единственные попытки придать эволюционный смысл гипотезе запрограммированного старения, но Алекс Ковальд и Томас Кирквуд подробно разобрали математические модели «программистов» и в каждой из них нашли серьезные ошибки и/или несоответствия.
Также встречается такой аргумент, что существование эпигенетических часов, которые тесно коррелируют с возрастом человека (и других млекопитающих), является доказательством наличия программы старения. Но это является ничем иным как логической ошибкой: в автомобиле, например, есть одометр, показания которого (пробег авто) тесно коррелируют с возрастом автомобиля, однако это вовсе не означает, что автомобиль начинает функционировать хуже, потому что с определенного пробега включается специальная программа. Частные случаи, впрочем, могут быть, но в общем, это не так.
Подытожим:
Гипотетическая программа старения вредит индивидуальному отбору и НЕ дает никаких ощутимых преимуществ при групповом отборе (который до сих пор не является общепризнанным).
Гипотеза запрограммированного старения нарушает принцип бритвы Оккама: постулируется новая сущность без надобности (см. пункт 1).
До сих пор не найдено ни намека на генетическую программу старения у млекопитающих хотя другие генетические программы давно найдены (например, вышеупомянутая программа отключения фертильности)
Самое главное: не существует примеров взломов «программы старения» внутри одного вида. Если есть программа, значит, чисто статистически должны быть особи, у которых она ломается со временем. Например, у каждой клеточки нашего организма есть программа самоубийства (апоптоза), но она с завидной регулярностью ломается, что служит одним из механизмов патогенеза онкологических заболеваний. Где среди 7 миллиардов людей нестареющие мутанты с отключенной программой старения?! Ведь ожидаемая продолжительность жизни таких людей должна составлять как минимум сотни лет.
С эволюционной точки зрения старение по-прежнему лучше всего описывается различными модификациями гипотезы Медавара, которая сводится к тому, что в дикой природе смертность от старения у большинства видов пренебрежимо мала в сравнении со смертностью от внешних причин (болезни, хищники и т. п.) и поэтому мутации, которые увеличивают максимальную продолжительность жизни в популяции не закрепляются. Однако те виды, которые не испытывают такого давления среды (очень крупные животные или живущие в защищенной от хищников среде), живут гораздо дольше. Дальнейшим развитием идеи Медавара является гипотеза антагонистической плейотропии, которая утверждает, что в условиях дикой природы эволюция закрепит те варианты генов, которые могут снижать максимальную продолжительность жизни, но увеличивают вероятность репродуктивного успеха в молодости. Гипотеза Медавара была не единожды проверена экспериментально:
В лабораторных условиях эволюционные гипотезы можно эффективно проверять с помощью экспериментальных эволюционных парадигм. Это подразумевает применение специфичного режима отбора и проведение мониторинга эволюционных изменений на протяжении поколений. Например, Стернс и коллеги на протяжении 60 поколений применяли к одной группе плодовых мушек высокую случайную смертность, а к другой группе — низкую случайную смертность. В результате оказалось, что режим высокой смертности приводил к укорочению продолжительности жизни, снижению возраста полового созревания и ускорению репродуктивной траектории по сравнению с режимом низкой смертности. Эти данные оказались полностью сопоставимыми с результатами природного эксперимента, в котором опоссумов, естественным образом эволюционирующих на лишенном хищников острове, сравнивали с опоссумами нормальной популяции на населенном хищниками материке.
Но это все эволюционные гипотезы, которые не описывают конкретные механизмы, а ознакомиться с наиболее правдоподобным на мой взгляд гипотетическим механизмом старения можно здесь.
