Фармакологическая модуляция памяти. Часть 1
После того, как в общих чертах освещены механизмы долговременной и кратковременной рабочей памяти, проследуем (наконец-то!) от нейрофизиологии к фармакологии и рассмотрим, какие вещества и каким способом могут оказывать позитивное влияние на память.
Предупреждение. Я категорически не советую употреблять ни одно из ниже перечисленных веществ без показаний и без назначения врача.
Ранее, в комментариях к моей заметке, посвящённой ноотропным препаратам, меня спрашивали, почему нет никаких доказательств того, что ноотропы имеют эффект, отличный от плацебо. В этой статье я представлю данные литературы, которые содержат такие доказательства. Будут даны ссылки на эксперименты, в которых память экспериментальных животных или людей специально не ослаблена физическим воздействием или химическими веществами.
Логика изложения будет такая. Сначала рассмотрим агенты, которые действуют на мембранные рецепторы, центры связывания которых находятся внеклеточно, т.е. на внешней стороне клеточной мембраны. Далее речь пойдёт о веществах, модулирующие внутриклеточные мишени. Для того, чтобы не загромождать публикацию, первая часть, посвящённая фармакологии мембранных рецепторов, будет в свою очередь разделена по типам рецепторов: глутаматные, дофаминовые, ацетилхолиновые. Хотя, конечно, есть и другие мишени. Может, дойду до них.
Итак, поехали.
Перед нами пустыня, почти голая, почти безжизненная синапс, образованный глутаматергическим, т.е. выделяющим глутамат, нейроном и нейроном, имеющим на своей поверхности ионоторопные, т.е. представляющие собой ионные каналы, глутаматные рецепторы: AMPA-рецепторы и NMDA-рецепторы. На рисунке ниже глутаматергическим является пресинаптический нейрон, а постсинаптический нейрон содержит глутаматные рецепторы.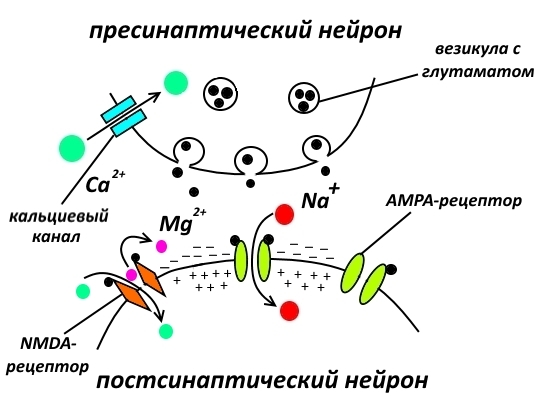
Те, кто внимательно читал два моих эссе о кратковременной и долговременной памяти, сейчас должен понять, что мы рассматриваем ситуацию долговременной потенциации и связанной с ней долговременной памятью.
Без работы AMPA- и NMDA-рецепторов невозможна передача нервного импульса и невозможно запоминание, поэтому логичным кажется, что стимулирование этих рецепторов с помощью соответствующих агонистов (т.е. веществ, которые запускают их работу) должно привести к улучшению памяти.
Но это не так. Прямое, я бы сказал «топорное», стимулирование рассматриваемых глутаматных рецепторов их агонистами может вызывать необратимое повреждение мозга и не приведёт к улучшению памяти.
Будем разбираться, почему.
Чрезмерная стимуляция NMDA- или AMPA-рецепторов приводит к избыточному поступлению в нейрон ионов кальция. Ионы кальция, как вы, наверное, уже поняли, являются активаторами многих внутриклеточных ферментов. Когда концентрация кальция возрастает выше определённых значений, активируются ферменты, ответственные за процесс апоптоза, т.е. клеточного самоубийства. Такая гибель нейронов под действием высоких концентраций возбуждающих нейромедиаторов носит название эксайтотоксичность (от англ. to excite — возбуждать, активировать).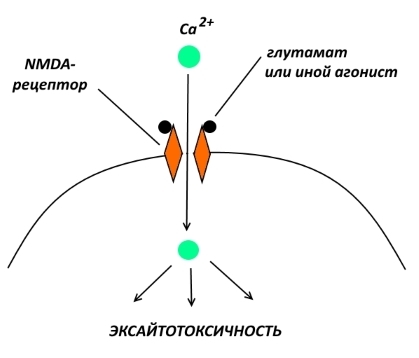
Примером вещества, вызывающего эксайтотоксичность, является домоевая кислота, выделяемая некоторыми видами морского планктона. По пищевой цепочке этот биотоксин может попадать в рыбу, а из неё — в организм других животных, в том числе и человека.
Домоевая кислота разрушает гиппокамп и миндалевидные тела, вызывая эксайтотоксичость посредством связывания с AMPA-рецепторами. Описано большое количество случаев отравления морских животных этим токсином. Есть мнение, что именно наблюдение за обдолб дезориентированными из-за токсина чайками привело к созданию культового фильма «Птицы».
Представим, однако, на минуту, что никакой эксайтотоксичности не существует. Возможно ли в позитивное влияние агонистов глутаматных рецепторов на память в таком случае?
Правильный ответ — едва ли.
Глутамат — основной возбуждающий медиатор в ЦНС. Каждые несколько миллисекунд миллионы нейронов в ЦНС генерируют потенциалы действия, возникающие посредством связывания глутамата с соответствующими рецепторами.
Вспомним, что долговременная потенциация возникает в том случае, когда один из нейронов резко увеличивает число сигналов (и/или их интенсивность) передаваемых другому. Именно такая ситуация происходит, например, при многократном повторении или при обучении. Но поступление глутамата в мозг приведёт к увеличению числа генериремых потенциалов действия ВСЕХ нейронов вне зависимости от их синаптических связей. Если бы это не приводило к негативным необратимым воздействиям, то усилилась бы связь всех нейронов со всеми, а не тех, которым это «надо» для решения той или иной когнитивной задачи.
Приведу такое сравнение. Радиожурналист берёт на стадионе интервью у спортсмена, который только что побил рекорд. Трибуны ликуют, болельщики кричат. Мы, чтобы услышать, что говорит новоиспечённый чемпион, увеличиваем громкость радиоприёмника. Но нам это не помогает, так как вместе со звуком голоса рекордсмена усиливаются и мешающие голоса трибун.
В качестве как будто бы исключения дам ссылку на статью [1], в которой внтрижелудочное введение глутаминовой кислоты привело к усилению памяти крыс. Статья, правда, опубликована в журнале, не вызывающем большого доверия, да в ней ко всему ещё и есть опечатки. Однако, если верить её авторам, эффет глутамата статистически отличен от плацебо и длится до 7 суток.
И всё же, несмотря на эксайтотоксичность, путь моделирования памяти посредством влияния на глутаматные рецепторы вовсе не заказан!
Более того, вещества, модулирующие активность глутаматных рецепторов, относятся к классу наиболее распространённых препаратов, улучшающих когнитивные функции, — к классу так называемых ампакинов (от названия AMPA-рецепторов), куда входят рацетамы, нахоящиеся у многих на слуху.
Каков же механизм их действия?
Как и многие биологические мишени, AMPA-рецепторы могут связываться не только с агонистами или антогонистами, но и с так называемыми аллостерическими модуляторами. Аллостерические модуляторы связываются с аллостерическим центром (от греч. «allos» — другой) — частью рецепторной молекулы, отличной от той её части, в которой происходит связывание с естественным лигандом.
Аналогия может быть такая. Детское поведение контролируется мозгом, расположенным в голове. Однако многие родители для изменения этого поведения находят весьма действенным воздействие на иную часть тела, лишённую большого количества нервным волокон, но богатую мышечной тканью. Таким образом, эту часть тела можно назвать аллостерическим центром детского организма по отношению к голове.
Важно понимать, что связывание той или ной молекулы (в наше случае — ампакина) с аллостерическим центром AMPA-рецептора не приводит к открытию ионного канала, поступлению внутрь нейрона положительных зарядов и генерации потенциала действия! Говорят, что позитивный аллостерический модулятор лишён внутренней активности. Все эти последствия могут вызвать только агонисты. Но в чём же тогда польза от аллостерических модуляторов? 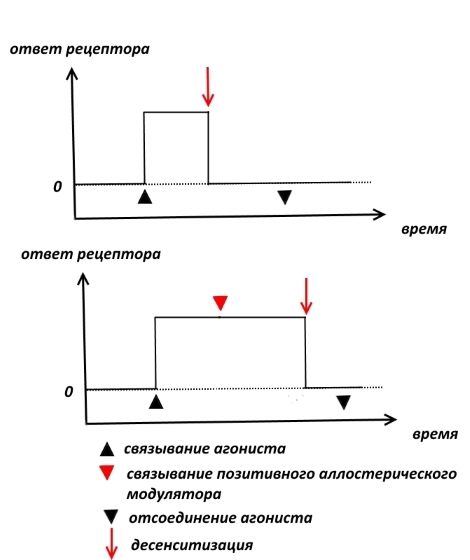
После пропускания порции положительно заряженных ионов, даже если глутамат всё ещё связан с AMPA-рецептором, происходит закрытие ионного канала и прекращение его работы. Такое явление называется десенситизацией (потеря чувствительности от лат. «sensus» — чувство, ощущение). Десенситизация призвана уберечь нервную систему от перевозбуждения. Позитивные аллостерические модуляторы AMPA-рецепторов замедляют наступление десенситизации, продлевая работу рецепторов. В результате долговременная потенциация усиливается, и памятный след вырабатывается быстрее, а затухает медленнее.
Замедление десенситизации связано с изменением конформации (формы) рецептора.
Позитивные аллостерические модуляторы AMPA-рецепторов класса рацетамов являются самыми «древними» ноотропными препаратами, а первый, наиболее известный их представитель, до сих пор продаваемый в Европе и странах бывшего СССР, — пирацетам.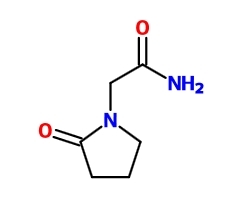
Влияние пирацетама на память здоровых людей изучалось неоднократно, в исследованиях были заметны позитивные отличия от плацебо, однако выборки были невелики [2], [3], [4], [5].
В перечисленных выше исследованиях изучали работу не только долговременной, но и оперативной памяти, а сейчас известно, что AMPA-рецепторы принимают в функционировании последней не самое важное участие. Вероятно, по этой причине отличия от плацебо хоть и значимые, но не очень сильные.
В клиническую практику внедрены аналоги пирацетама (в настоящее время используются не все из них): прамирацетам, анирацетам, оксирацетам, фонтурацетам, однако, к сожалению, их влияние на неугнетённую память людей в литературе не описано, в то время как примеров позитивного влияния на память здоровых животных — уйма. Я ограничусь следующими обзорами: [6], [7], [8].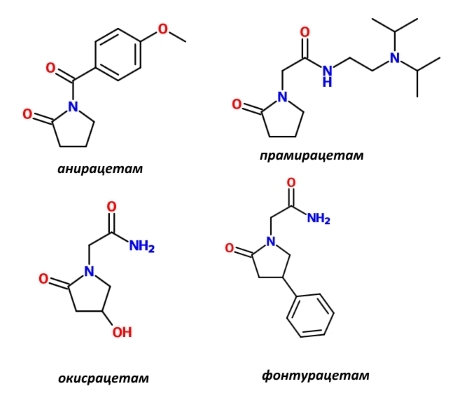
Особняком стоит отметить ещё один рацетам — леветирацетам. В отличие от других рацетамов, призванных улучшать состояние когнитивной сферы при её различного рода нарушениях, леветирацетам предназначен для лечения эпилепсии. Его позитивное воздействие на здоровых людей было изучено [9].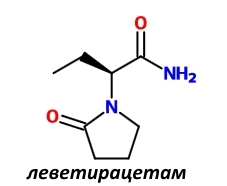
Отмечу, что до сих пор фармакологи не теряют надежды получить значимый позитивный эффект от этой группы препаратов при лечении нейродегенативных и психических заболеваний [10]. Среди последних попыток можно назвать CX516 и фарампатор, у которых отмечались позитивные эффекты (очень небольшие) в отношении когнитивной сферы здоровых добровольцев [11], [12].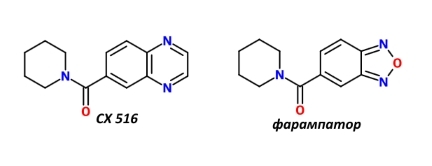
Продолжение следует.
Библиография.
1.Тюренков И.Н., Багметова В.В., Чернышева Ю.В., Меркушенкова О.В., Берестовицкая В.М., Васильева О.С. Сравнение психотропных свойств глутаминовой кислоты и её нового производного — гидрохлорида бета-фенилглутаминовой кислоты (глутарона)// Фундаментальные исследования. 2013, № 3. С. 167–172.
2.S.J. Dimond, E.Y. M. Brouwers. Increase in the power of human memory in normal man through the use of drugs// Phychopharmacology. 1976, vol. 49, issue 3. P. 307–309.
3.C. Wilsher, G. Atkins, P. Manfield. Piracetam as an aid to learning in dyslexia// Phychopharmacology. 1979, vol. 65, issue 1. P. 107–109.
4.P. Mindus, B. Cronholm, S.E. Levander, D. Schalling. Piracetam-induced improvement of mental performance// Acta Psychiatrica Scandinavica. 1976, vol. 54, issue 2. P. 150–160.
5.H.M. Alkuraishy, A.I. Algareeb, A.K. Albuhadily, B.M. Almgoter. Modulation effects of piracetam ang Ginkgo biloba on the cognitive and working memory functions: psychometric study// Neurology and Neurophysiology. 2014, vol. 5, issue 5. doi: 10.4172/2155–9562.1000234
6.A.H. Gouliaev, A. Senning. Piracetam and structurally related nootropics// Brain Research Reviews. 1994, vol. 19, issue 2. P. 180–222.
7.A.G. Malykh, M.R. Sadaie. Piracetam and piracetam-like drugs. From basic science to novel clinical application to CNS disorders// Drugs. 2010, vol. 70, issue 3. P. 287–312.
8.F. Gualtieri, D. Manetti, M.N. Romanelli, C. Ghelardini. Design and study of piracetam-like nootropics, controversial members of the problematic class of cognition-enhancing drugs// Current Pharmaceutical Design. 2002, vol. 8, issue 2. P. 125–138.
9.J.C. Magalhães, M. Gongora, R. Vicente, J. Bittencourt, G. Tanaka, B. Velasques, S. Teixeira, G. Morato, L.F. Basile, O. Arias-Carrión, F.A. M.S. Pompeu, M. Cagy, P. Ribeiro. The influence of levetiracetam in cognitive performance in healthy individuals: neuropsychological, behavioral and electrophysiological approach// Clinical Psychopharmacology and Neuroscience. 2015, vol. 13, issue 1. P. 83–93.
10.В.В. Григорьев, А.Н. Прошин, А.С. Кинзирский, С.О. Бачурин. Современные подходы к созданию симуляторов памяти и когнитивных функций на основе лигандов AMPA-рецепторов// Успехи химии. 2009, том 78, № 5. С. 524–534.
11.M. Ingvar, J. Ambros-Ingerson, M. Davis, R. Granger, M. Kessler, G.A. Rogers, R.S. Schehr, G. Lynch. Enhancement by an ampakine of memory encoding in humans// Experimental Neurology. 1997, vol. 146, issue 2. P. 553–559.
12.E. Wezenberg, R.J. Verkes, G.S. F. Ruigt, W. Hulstijn, B.G. C. Sabbe. Acute effects of the ampakine farampator on memory and information processing in healthy eldery volunteers// Neuropsychopharmacology. 2007, vol. 32, issue 6. P. 1272–1283.
